常见细胞污染类型如何辨别及预防解决方法
常见细胞污染类型如何辨别及预防解决方法:细胞培养中常见的生物污染类型有7种,分别是细菌污染,支原体污染,原虫污染,黑胶虫污染,真菌污染,病毒污染以及非细胞污染,真菌污染来源,一般是来自实验服,并且具有气候性,多雨······

发布时间:2026-01-21 17:30:00 细胞资源库平台 访问量:220
乙肝病毒(HBV)感染是全球主要公共卫生问题之一,有超过2.5亿人慢性感染HBV。而其中有超三分之一的人口集中在我国,人数接近1亿人。NTCP工具细胞,特别是外源表达NTCP的肝癌细胞系如HepG2-NTCP和Huh7-NTCP,因其易操作、短周期、重现性佳的特点,在乙肝病毒(HBV)研究中扮演着至关重要的角色。这些细胞模型能够有效模拟HBV的感染过程,为研究HBV的生命周期、宿主限制因子、病毒复制以及药物筛选提供了一个强大而便捷的体外平台。它们不仅有助于揭示HBV感染的分子机制,如DDX3作为宿主限制因子阻碍cccDNA转录,GPC5作为附着因子在感染入胞过程中的作用,还能通过直接与NTCP相互作用或下调NTCP表达来筛选和验证抗病毒药物的活性,例如环孢菌素A及其衍生物、雷帕霉素及其衍生物等。此外,这些工具细胞还促进了对HBV宿主特异性分子的发现,为发展支持HBV感染的小动物模型提供了可能,这对于乙肝相关研究和药物开发具有重大意义。
英文标题:N-terminal PreS1 Sequence Regulates Efficient Infection of Cell Culture-generated Hepatitis B Virus
中文标题:N 端 preS1 序列调控细胞培养衍生乙肝病毒(HBVcc)的高效感染
发表期刊:《Hepatology》
影响因子:15.8
作者单位:
1.Department of Virology II, National Institute of Infectious Diseases, Tokyo, Japan
2.Department of Gastroenterology and Hepatology, Shin-Yurigaoka General Hospital, Kawasaki, Japan
3.Liver Cancer Prevention Research Unit, Center for Integrative Medical Sciences, RIKEN, Wako, Japan
4.Department of Microbiology, Graduate School of Medicine, University of Yamanashi, Yamanashi, Japan
5.Department of Virology and Parasitology, Hamamatsu University School of Medicine, Shizuoka, Japan
6.Genome Medical Sciences Project, National Center for Global Health and Medicine, Ichikawa, Japan
7.Liver Diseases Branch, National Institute of Diabetes and Digestive and Kidney Diseases, National Institutes of Health, Bethesda, MD, USA
作者信息:
Asako Murayama,Norie Yamada
HBV 细胞培养系统是病毒特性研究和抗病毒药物开发的关键工具,但现有系统存在局限:HepG2.2.15/HepAD-38 等整合型细胞系来源的病毒,无法用于研究病毒多态性或耐药突变;瞬时转染普通 HBV 分子克隆获得的 HBVcc(细胞培养衍生 HBV)感染效率极低。研究发现 HBV 基因型 D(GTD)preS1 区存在 11 个氨基酸缺失(d11),而基因型 C(GTC)无此缺失,且 GTD 的 HBVcc 感染效率更高。本研究旨在验证 preS1 区 d11 对 HBVcc 感染效率的调控作用,明确其分子机制,建立高效 HBVcc 感染系统。
构建含 1.38 倍基因组的多基因型 HBV 克隆(A、B、C、D 型),通过瞬时转染 HepG2 细胞制备 HBVcc,在 preS1 区引入 d11 突变(如 GTC-d11)。以 HepG2/NTCP 细胞、PXB 人源肝细胞为感染模型,检测指标包括:1)HBc 免疫荧光染色计数感染细胞;2)RT-qPCR 检测 pre-genomic RNA(pgRNA);3)Hirt 提取法结合 qPCR 检测 cccDNA;4)碘克沙醇密度梯度离心分析病毒颗粒特性;5)Western blot 检测 HBs 蛋白(L/S-HBs)及其糖基化水平;6)HBV/NL 报告病毒系统、丁型肝炎病毒(HDV)包装系统验证关键分子(L-HBs、HBp、病毒基因组)的作用;7)细胞附着 / 内化实验探究感染机制。
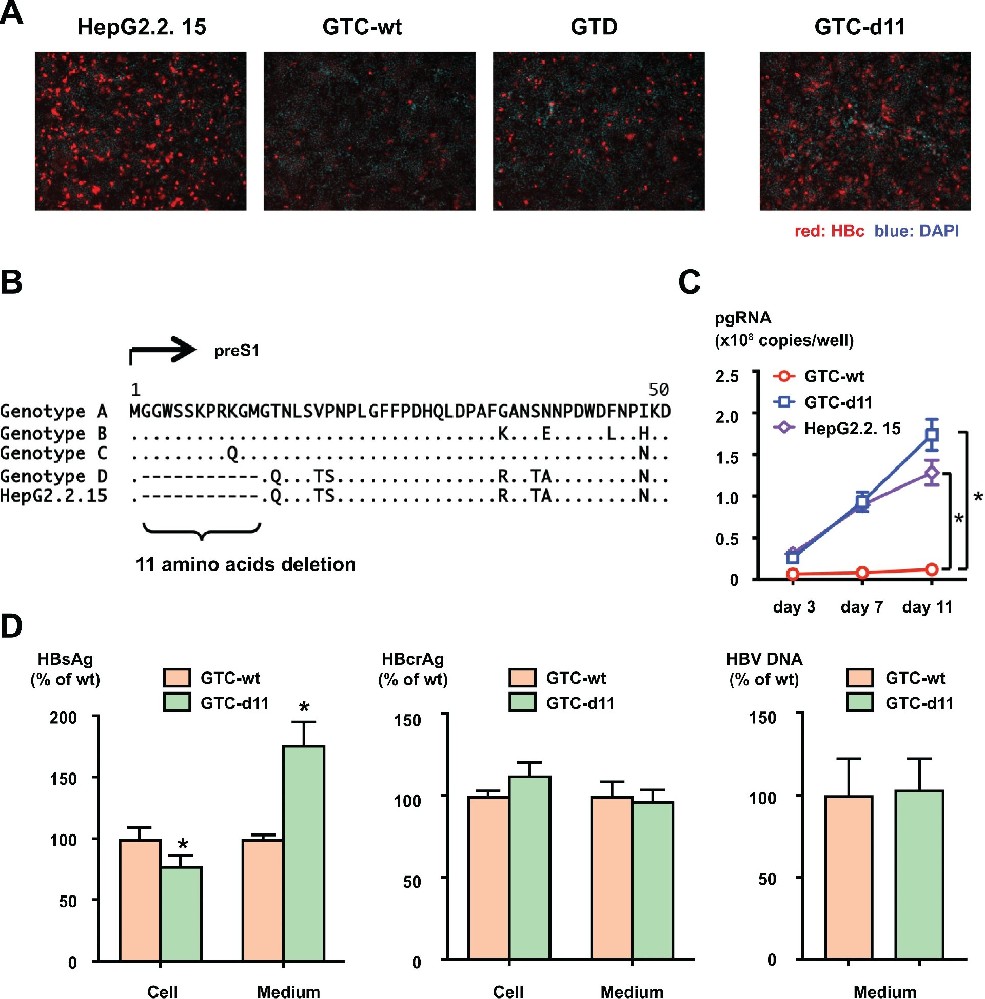
图 1:preS1 区 d11 增强 HBVcc 的感染效率
GTC-wt(野生型基因型 C)HBVcc 感染 HepG2/NTCP 细胞后,HBc 阳性细胞极少;引入 d11 后(GTC-d11),感染细胞数显著增加,与 HepG2.2.15 来源病毒(基因型 D)相当(图 1A)。序列比对显示,基因型 D 天然存在 preS1 区 11 个氨基酸缺失(图 1B)。GTC-d11 感染后,pgRNA 呈时间依赖性升高,而 GTC-wt 无此现象(图 1C);转染细胞的 HBsAg 分泌量更高,HBcrAg 和 HBV DNA 水平与 GTC-wt 无差异,表明 d11 不影响病毒复制,仅提升感染效率(图 1D)。
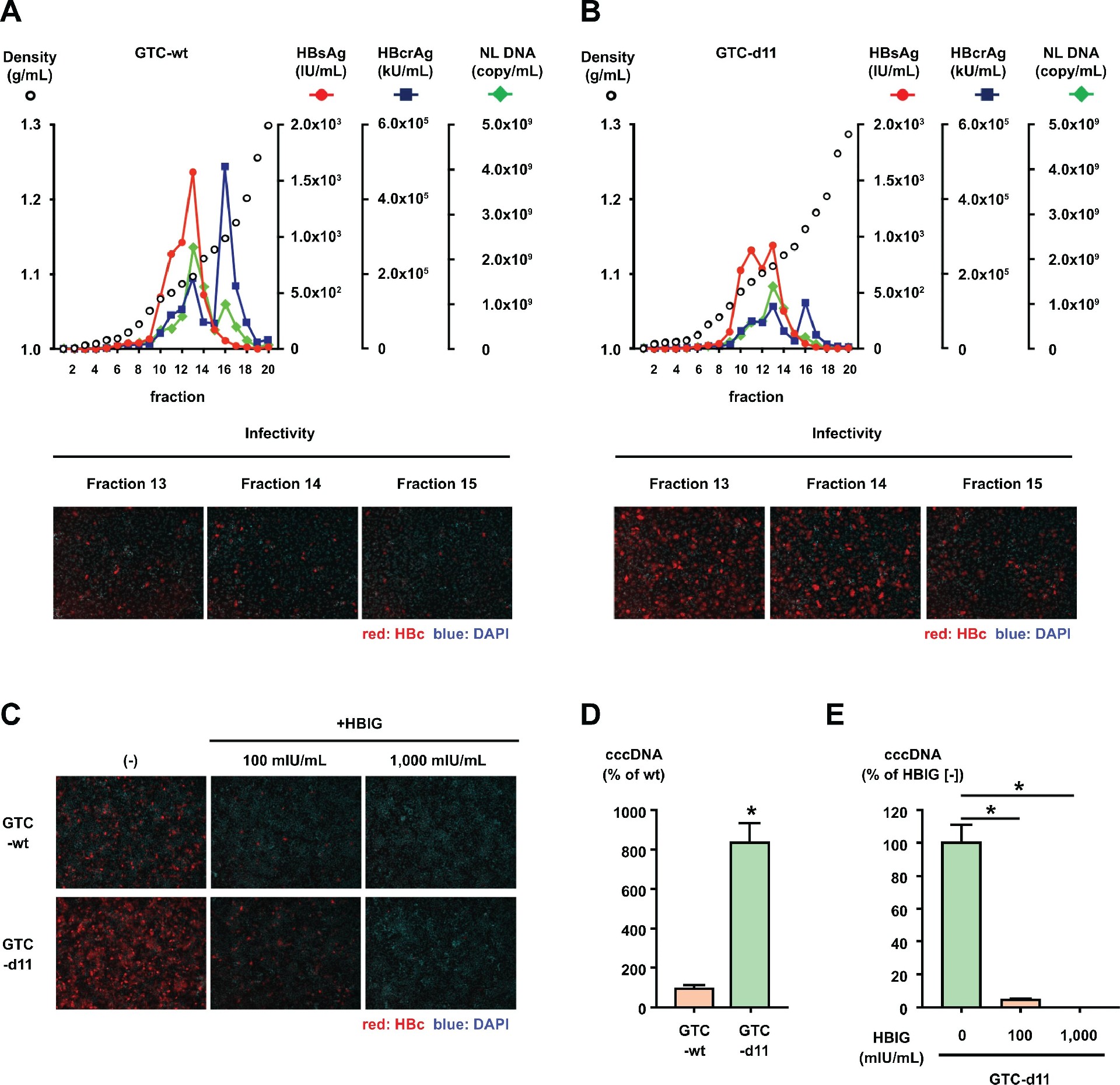
图 2:HBVcc 的密度梯度分析与 cccDNA 合成
碘克沙醇密度梯度离心显示,GTC-wt 和 GTC-d11 的 HBsAg、HBcrAg、HBV DNA 及感染性峰值均位于相同组分(图 2A、B),但相同 HBV DNA 剂量下,GTC-d11 的感染效率显著更高,且感染可被 HBIG 抑制(图 2C)。GTC-d11 感染细胞的 cccDNA 合成量是 GTC-wt 的 8.5 倍,且 HBIG 可阻断该过程(图 2D、E),证实 d11 增强的是依赖 HBs 的特异性感染。
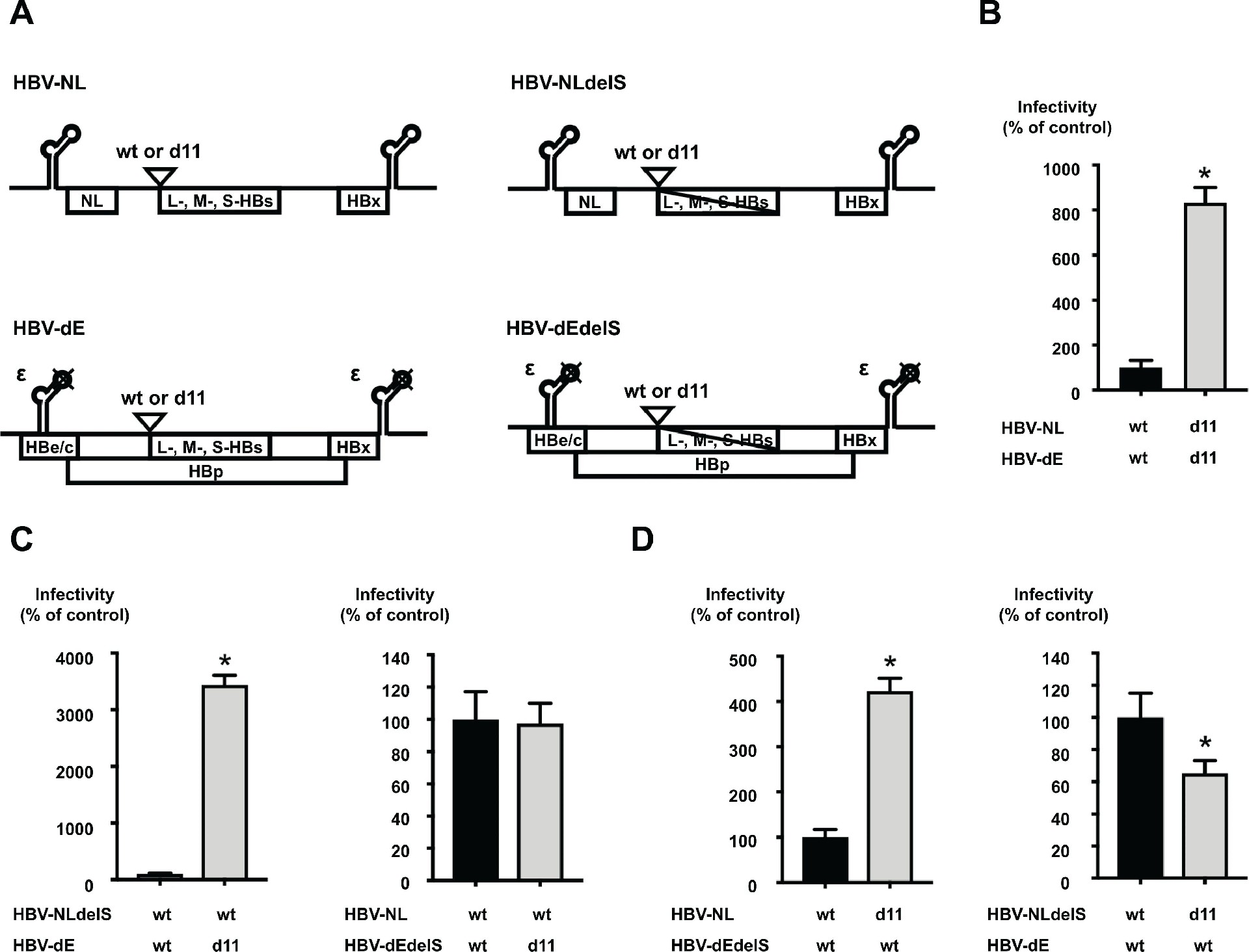
图 3:L-HBs 蛋白的 d11 是感染增强的关键
HBV/NL 报告病毒系统中,d11 引入后(HBV-NL-d11+HBV-dE-d11),荧光素酶活性提升 8.3 倍(图 3B)。通过缺失 HBs 表达的突变体验证:仅当 L-HBs 含 d11 时感染增强,HBp 含 d11 或病毒基因组缩短无此效果(图 3C、D);逐步缺失实验显示,仅 11 个氨基酸完整缺失(d11)可实现最大感染增强(补充图 S3),证实 L-HBs 的 d11 是核心调控因素。
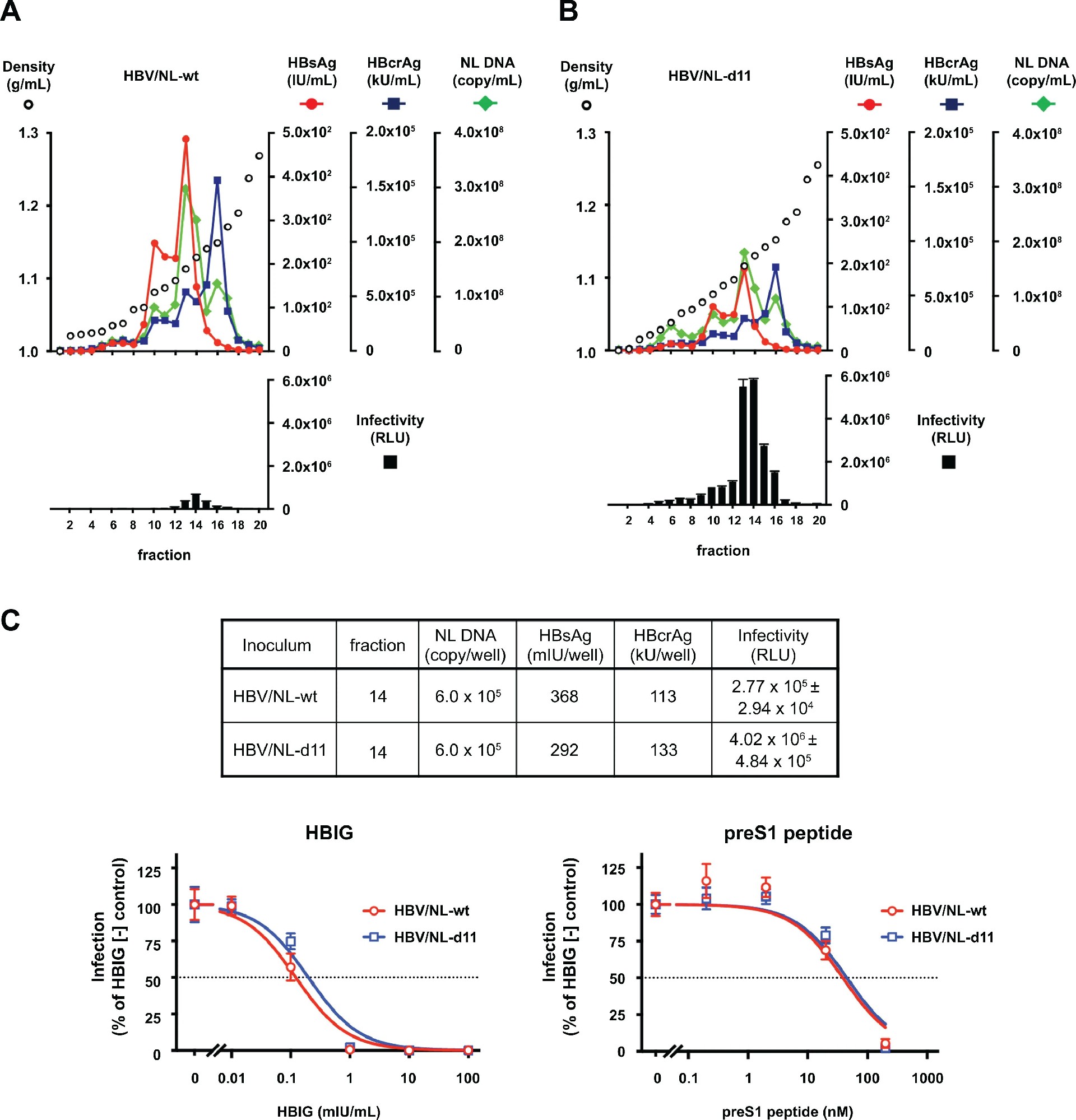
图 4:HBV/NL 报告病毒的特性与药物敏感性
HBV/NL-d11 的密度梯度分布与野生型一致,但相同 NL DNA 剂量下,感染效率提升 14.5 倍(图 4A、B、C 上)。HBIG 和 preS1 肽(Myrcludex B)可剂量依赖性抑制两种病毒感染,且 EC₅₀值无差异(图 4C 下),表明 d11 不改变病毒对 HBs 靶向抑制剂的敏感性。
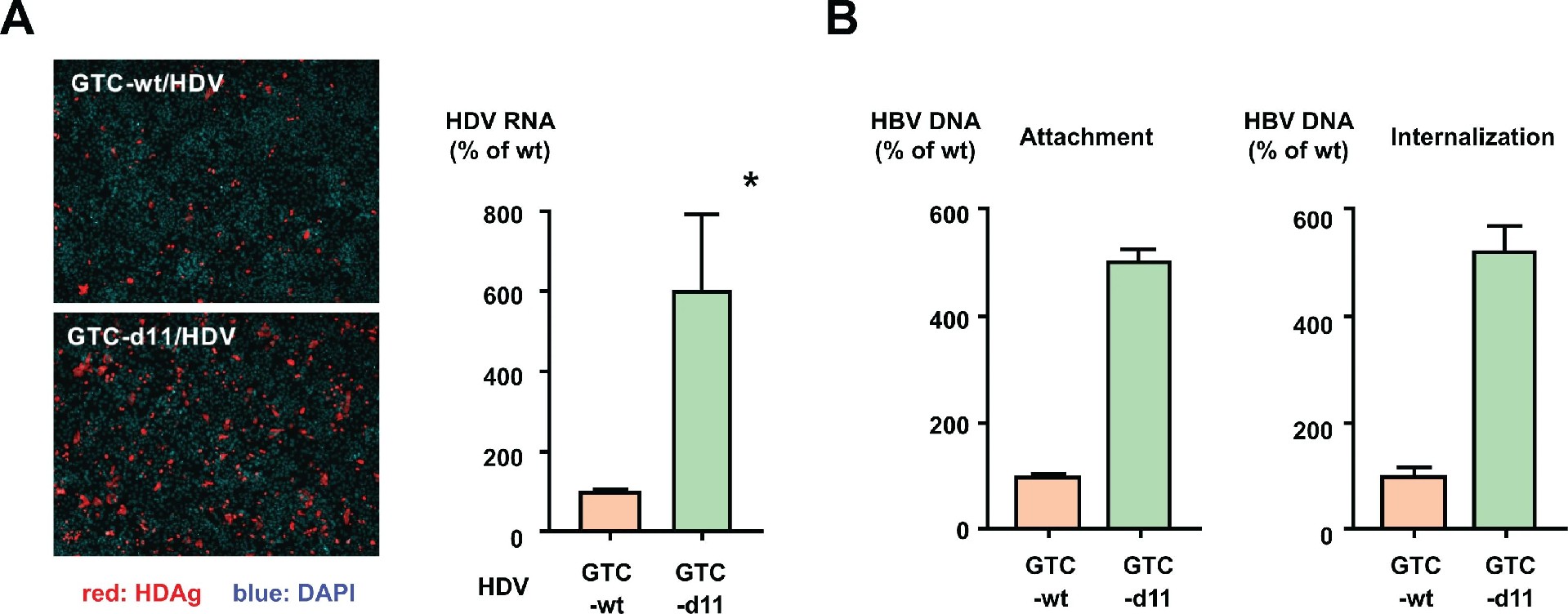
图 5:d11 增强病毒细胞附着并提升 L-HBs 糖基化
HDV 包装实验显示,含 d11-L-HBs 的 HDV 感染效率是野生型的 6 倍(图 5A),证实 L-HBs 的 d11 可跨病毒增强感染。细胞附着实验显示,4℃下 GTC-d11 在细胞表面的附着量是 GTC-wt 的 5.1 倍,内化量同步升高(图 5B);即使在无 NTCP 的 HepG2 细胞上,GTC-d11 的附着量仍高 2 倍,提示 d11 增强病毒与细胞表面分子(如硫酸乙酰肝素蛋白聚糖)的相互作用。Western blot 显示,GTC-d11 的 L-HBs 糖基化比例(64.7%-68.7%)显著高于 GTC-wt(33.5%-42.9%),且与高感染性的 HepG2.2.15 病毒一致,而 S-HBs 糖基化无差异(补充图 S5)。
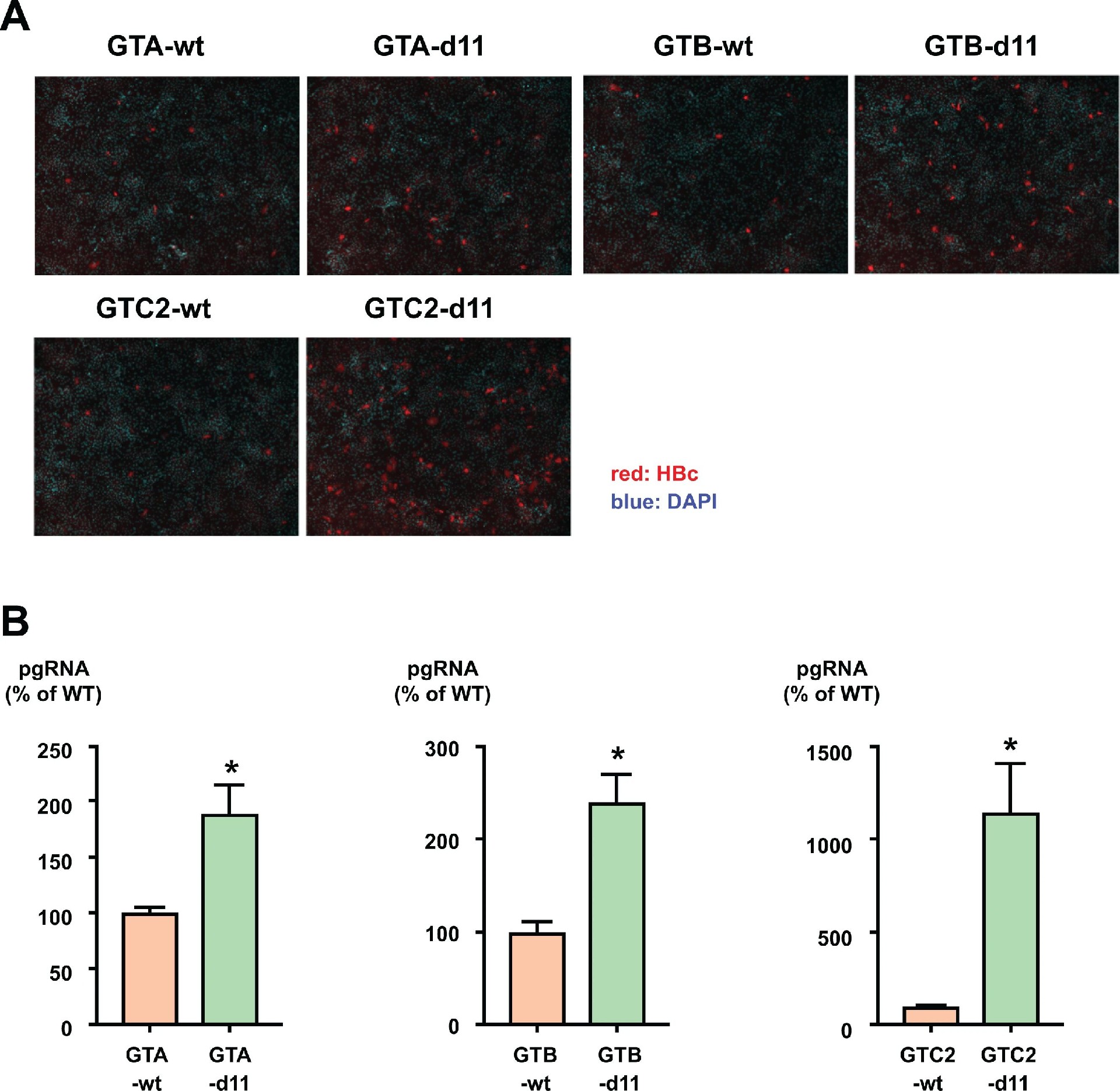
图 6:d11 的感染增强效应适用于多基因型 HBV
在基因型 A(GTA)、B(GTB)及另一基因型 C 克隆(GTC2)中引入 d11 后,HBc 阳性细胞数均显著增加(图 6A);pgRNA 水平分别提升 1.7 倍(GTA-d11)、2.4 倍(GTB-d11)和 11 倍(GTC2-d11)(图 6B),证实 d11 的感染增强效应不依赖 HBV 基因型或菌株。
本研究明确 preS1 区 11 个氨基酸缺失(d11)是调控 HBVcc 高效感染的关键因素,核心机制如下:1)d11 通过 L-HBs 蛋白发挥作用,与 HBp 或病毒基因组长度无关;2)d11 增强病毒与细胞表面的附着能力(包括 NTCP 依赖和非依赖途径);3)d11 提升 L-HBs 的糖基化水平,可能优化病毒包膜结构与细胞的相互作用。该突变可显著提升多基因型 HBVcc 的感染效率,且不改变病毒对 HBs 靶向抑制剂的敏感性,同时支持高效 cccDNA 合成。该系统解决了传统 HBVcc 感染效率低的痛点,为 HBV 多态性研究、耐药突变分析及抗病毒药物(尤其是靶向感染步骤或 cccDNA 合成的药物)筛选提供了高效工具。
常见细胞污染类型如何辨别及预防解决方法:细胞培养中常见的生物污染类型有7种,分别是细菌污染,支原体污染,原虫污染,黑胶虫污染,真菌污染,病毒污染以及非细胞污染,真菌污染来源,一般是来自实验服,并且具有气候性,多雨······
细胞聚团的原因分析及如何避免:培养物中细胞可能聚集的一些原因包括:1.过度消化、2.环境压力、3.组织分解、4.过度生长、5.污染等;如何避免聚团细胞的生成;首先确认当前细胞生长密度及状态,80%左右的生长密度即可进行······
细胞有空泡原因分析及解决方法:出现细胞空泡情况有1.细胞老化2.培养液错误配制;3.细胞消化时操作不当;4.污染等等,如细胞老化,解决方法,原代细胞使用较低代次进行实验,传代细胞避免传代次数过高···
细胞半换液和全换液操作步骤:第一种方式:细胞全换液;如果是贴壁细胞,可以用全量换液法,直接吸去全部旧培养基,补充足量新鲜完全培养基;第二种方式:细胞半换液;"细胞半换液"又称"细胞半量换液",即弃掉一半旧的培养基,再······
细胞生长缓慢的可能原因有哪些:细胞培养外部因素包括细胞培养基的配方和质量问题,培养条件不理想,污染问题,细胞自身因素包含细胞的健康状态,细胞密度过高或过低,细胞老化现象,细胞特性,当细胞生长出现缓慢的问题时,我······
常用胰腺癌细胞株动物模型及胰腺癌细胞株有哪些:胰腺癌研究中常用的动物模型主要包括化学物质诱导胰腺癌动物模型,基因工程胰腺癌小鼠模型和胰腺癌移植模型,常用的胰腺细胞株MIA-PACA-2人胰腺癌细胞,Capan-2人胰腺癌细······
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
上一篇:IκBβ/NFκB 信号通路缺失不能减轻对乙酰氨基酚诱导的肝损伤
下一篇:上皮细胞分泌的细胞外基质 niche 驱动肠道类器官形成研究
厦门爱恪信生物科技有限公司
手机:15859239971
邮箱:2205839769@qq.com
地址:厦门翔安火炬高新区翔星路96号建业楼D座602

微信公众号
ATCC细胞培养

技术支持
15859239971
Copyright©厦门爱恪信 闽ICP备19027235号-7
公安备案: XML地图
XML地图




 客服QQ
客服QQ