常见细胞污染类型如何辨别及预防解决方法
常见细胞污染类型如何辨别及预防解决方法:细胞培养中常见的生物污染类型有7种,分别是细菌污染,支原体污染,原虫污染,黑胶虫污染,真菌污染,病毒污染以及非细胞污染,真菌污染来源,一般是来自实验服,并且具有气候性,多雨······

发布时间:2025-12-11 09:00:00 细胞资源库平台 访问量:187
Reporter THP1 Cell Line 细胞系是以 THP-1 为工具细胞,采用慢病毒感染的方式,构建稳定表达报告基因的细胞系,可用于检测信号通路转导。在被PMA、INF-α等刺激后,目标信号通路被激活,会启动下游特定基因的表达,这个表达过程会同时带动“报告基因”的表达。通过检测报告基因的信号强度,可以间接、定量、灵敏地衡量该信号通路的激活程度。
THP-1细胞本身来源于人单核细胞,可以分化为巨噬细胞样细胞,是研究先天免疫的经典模型。报告基因株的建立使其功能更强大。THP-1报告基因株是免疫学、炎症性疾病、药物研发等领域不可或缺的强大工具。它可以将一个复杂的、内在的生物学过程(信号通路激活/基因表达)转变为一个易于检测、可定量的物理信号(光或荧光)。这使得研究人员能够高通量地筛选药物,精确地量化免疫反应强度,直观地研究信号通路机制。
英文标题:High-Throughput Screening for CEBPD-Modulating Compounds in THP-1-Derived Reporter Macrophages Identifies Anti-Inflammatory HDAC and BET Inhibitors
中文标题:高通量筛选鉴定 CEBPD 调控化合物:抗炎 HDAC 与 BET 抑制剂的发现
发表期刊:《Internationaljournal of Molecular Sciences》
影响因子:4.9
作者单位:
1.Fraunhofer Institute for Translational Medicine and Pharmacology ITMP, Theodor-Stern-Kai 7, 60596 Frankfurt am Main, Germany
2.Fraunhofer Institute for Translational Medicine and Pharmacology ITMP, Schnackenburgallee 114, 22525 Hamburg, Germany
3.EpiEndo Pharmaceuticals ehf, Eiðistorg 13-15, 170 Seltjarnarnes, Iceland
作者信息:
Tatjana Ullmann¹*, Sonja Luckhardt¹, Markus Wolf², Michael J. Parnham¹,³, Eduard Resch¹*
CCAAT / 增强子结合蛋白 δ(CEBPD)是巨噬细胞炎症反应的核心转录因子,通过调控炎症基因转录参与类风湿关节炎、动脉粥样硬化等疾病进程。现有抗炎药物(如甾体、非甾体抗炎药)存在胃肠道损伤、免疫抑制等副作用,亟需开发靶向 CEBPD 的新型精准抗炎化合物。本研究构建 THP-1 来源的 CEBPD 报告基因巨噬细胞模型,高通量筛选化合物库,鉴定调控 CEBPD 活性的表观遗传抑制剂,为炎症驱动疾病提供新治疗方向。
本研究首先通过病毒转导技术构建 THP-1 来源的 CEBPD::SEAP 报告基因细胞系,将分泌型碱性磷酸酶(SEAP)基因置于 CEBPD 启动子调控下,以 SEAP 活性反映 CEBPD 转录活性;随后采用包含 1280 个药理活性化合物的 LOPAC®1280 库和 774 个 FDA 批准药物的 ENZO®774 库,建立 “化合物预处理 1 小时→LPS(0.1μg/mL)+IFN-γ(20ng/mL)刺激 24 小时→检测 SEAP 酶活性” 的高通量筛选流程,同时设置 M0 对照组(未刺激)、M1 溶剂对照组(DMSO)及 M1 TSA 对照组(已知 HDAC 抑制剂)以排除干扰;通过 RT-qPCR 检测化合物对报告基因 CEBPD::SEAP、内源性 CEBPD 及炎症相关基因(IL-6、CCL2、IL-1β)mRNA 表达的影响,利用 CellTiter-Glo® 实验验证细胞活力以排除毒性干扰,借助 Western blot 初步探索 PI3K-AKT/TGF-β 信号通路在化合物调控 CEBPD 中的潜在作用,最终通过两次重复筛选确认 hit 化合物的可靠性。

图 1:SEAP 报告基因检测系统的性能验证
该图确认检测系统的可靠性:HEK293T 细胞中,CMV 启动子驱动的 SEAP 可稳定分泌,其酶活性在 10⁴倍稀释范围内呈严格线性关系(R²=0.99),覆盖从低到高的广泛检测范围(图 1b);Z’因子达 0.7(理想值≥0.5), intra-assay CV(批内变异)<10%、inter-assay CV(批间变异)<5%(表 1),表明系统灵敏度高、重复性好,可满足高通量筛选对稳定性和准确性的要求(图 1c)。
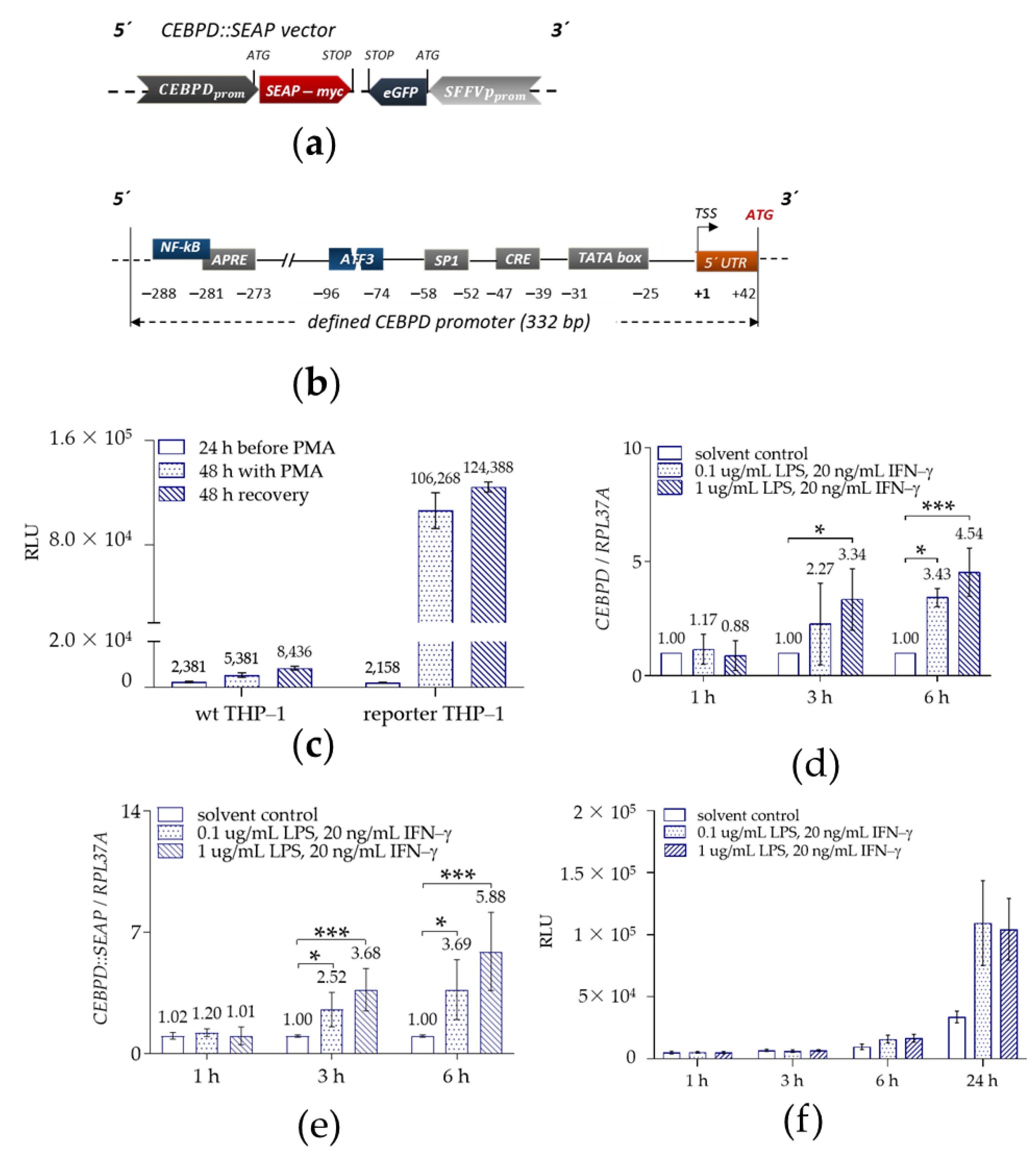
图 2:THP-1::CEBPD::SEAP 报告基因巨噬细胞系的有效性验证
该图证实细胞系能准确反映 CEBPD 活性:PMA(50ng/mL)处理 48h 可诱导 THP-1 报告细胞分化为贴壁巨噬细胞,分化过程中 CEBPD 启动子激活,SEAP 分泌量显著升高(图 2c);LPS(0.1/1μg/mL)+IFN-γ(20ng/mL)刺激可剂量依赖性上调内源性 CEBPD mRNA 表达,6h 时达峰值(最高 4.5 倍)(图 2d);报告基因 CEBPD::SEAP 的 mRNA 表达趋势与内源性 CEBPD 完全一致,且 SEAP 酶活性在刺激后 24h 达最高值(图 2e-f),说明该细胞系可精准模拟内源性 CEBPD 的转录调控,适合筛选 CEBPD 调控化合物。

图 3:高通量筛选流程及 hit 化合物鉴定
该图展示筛选策略与核心结果:建立标准化高通量筛选方案(图 3a),将化合物分为 “M0 对照组(未刺激)、M1 溶剂对照组(DMSO)、M1 TSA 对照组(已知 HDAC 抑制剂)、化合物处理组”,通过两次重复筛选排除假阳性;以 “SEAP 活性高于 M1 溶剂对照组 3 个标准差(3×SD)” 为 hit 判定标准,从 2054 个化合物中鉴定出 3 个核心 hit(I-BET151、Ro 11-1464、SAHA),并纳入已知 HDAC 抑制剂 TSA(图 3b-c);细胞活力实验显示,这些化合物的 SEAP 活性提升与细胞增殖 / 存活无关,排除细胞毒性或促增殖作用对结果的干扰。
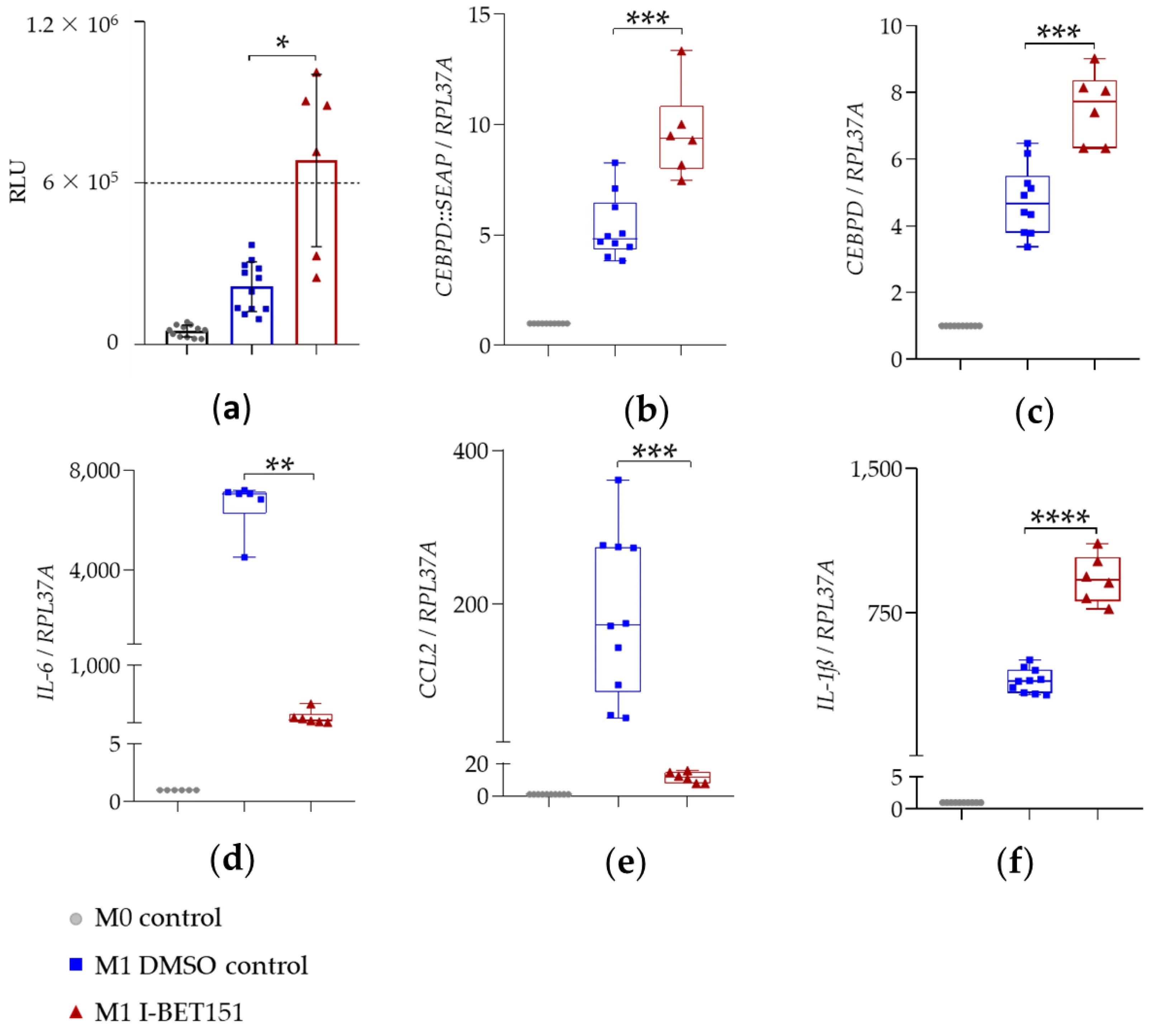
图 4:BET 抑制剂 I-BET151 对 SEAP 分泌及基因表达的影响
该图聚焦 I-BET151 的调控效应:10μM I-BET151 预处理可显著提升 LPS+IFN-γ 刺激后的 SEAP 酶活性(图 4a);基因表达层面,I-BET151 显著上调报告基因 CEBPD::SEAP(9.6 倍)和内源性 CEBPD(7.5 倍)的 mRNA 表达(图 4b-c、表 2);抗炎效果上,I-BET151 强效抑制 IL-6(从 6625 倍降至 102.3 倍,抑制 65 倍)和 CCL2(从 187.7 倍降至 11.5 倍,抑制 15 倍)的 mRNA 表达(图 4d-e、表 2);但同时上调 IL-1β 表达(从 396 倍升至 927.1 倍,增加 2.3 倍),显示其兼具抗炎与轻微促炎的双向作用(图 4f、表 2)。
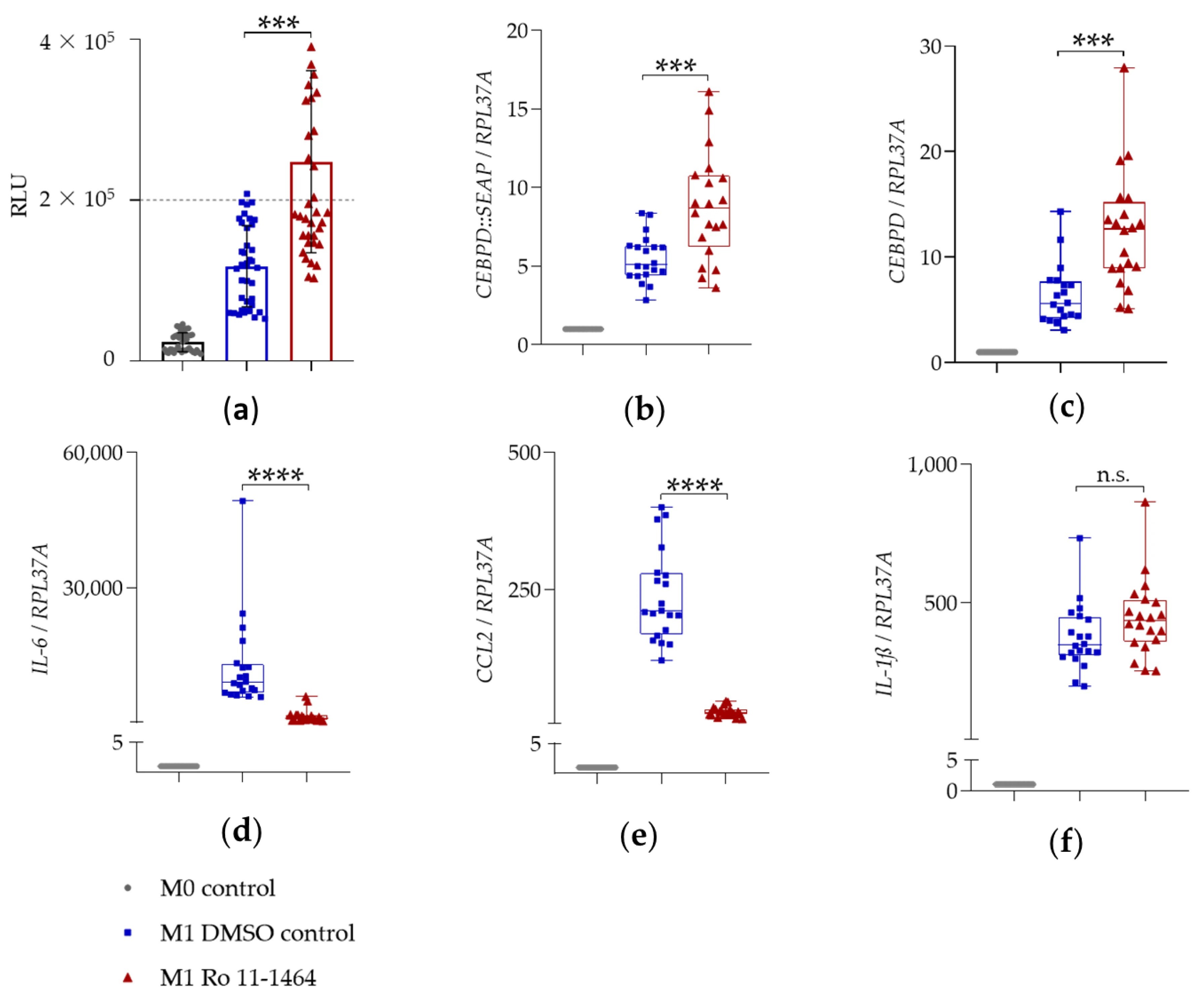
图5:BET 抑制剂 Ro 11-1464 对 SEAP 分泌及基因表达的影响
该图揭示Ro 11-1464 的特异性调控:10μM Ro 11-1464 预处理可显著升高 SEAP 酶活性,且效果具有统计学意义(p<0.001)(图 5a);与 I-BET151 类似,Ro 11-1464 可上调报告基因 CEBPD::SEAP(8.8 倍)和内源性 CEBPD(12.4 倍)的 mRNA 表达(图 5b-c、表 3);抗炎方面,Ro 11-1464 抑制 IL-6(从 12432 倍降至 1364 倍,抑制 9 倍)和 CCL2(从 237.2 倍降至 26.6 倍,抑制 9 倍)表达(图 5d-e、表 3);但与 I-BET151 不同,Ro 11-1464 对 IL-1β mRNA 表达无显著影响(344.8 倍 vs 445.3 倍,p>0.05),提示不同 BET 抑制剂对 IL-1β 的调控存在差异(图 5f、表 3)。
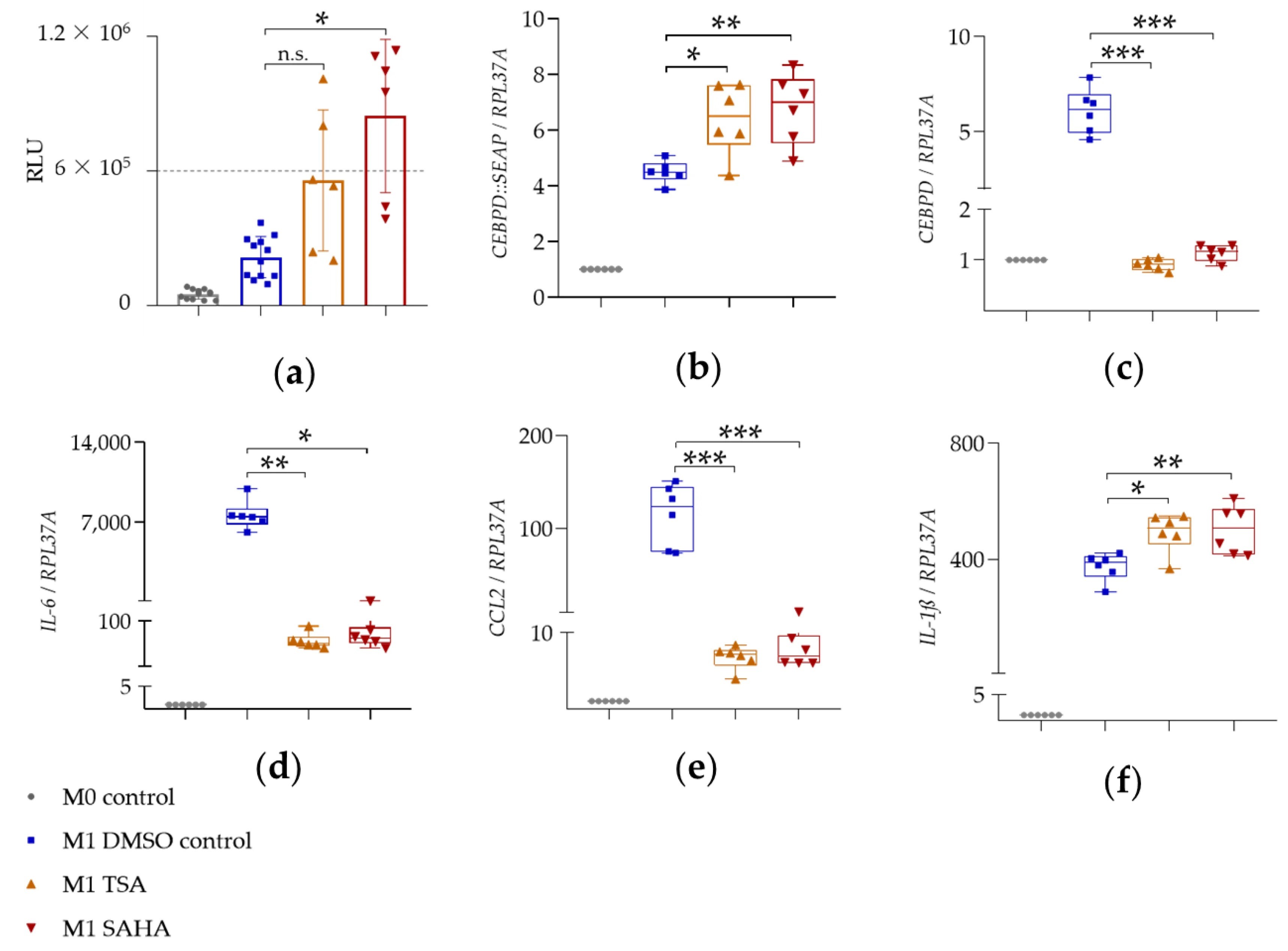
图 6:HDAC 抑制剂 SAHA 和 TSA 对 SEAP 分泌及基因表达的影响
该图展现 HDAC 抑制剂的独特调控模式:10μM SAHA 或 0.5μM TSA 预处理后,SEAP 酶活性显著升高(图 6a);但基因表达呈现 “双向差异”—— 报告基因 CEBPD::SEAP 的 mRNA 表达显著上调(SAHA 组 6.4 倍,TSA 组 6.8 倍),而内源性 CEBPD 的 mRNA 表达被完全抑制(接近 M0 对照组水平,表 4)(图 6b-c);抗炎效果上,SAHA 和 TSA 均强效抑制 IL-6(SAHA 抑制 110 倍,TSA 抑制 130 倍)和 CCL2(均抑制 15 倍以上)表达(图 6d-e、表 4);同时轻微上调 IL-1β 表达(SAHA 组 492.5 倍,TSA 组 501.9 倍),但幅度低于 I-BET151(图 6f、表 4),这种 “报告基因与内源性 CEBPD 调控相反” 的现象,推测与 HDAC 抑制剂对 CEBPD 启动子的顺式激活(报告基因仅含近端 332bp 启动子)和反式抑制(内源性基因含远端调控区域)的平衡差异有关
本研究通过高通量筛选从 2054 个化合物中鉴定出 4 种可调控 CEBPD 活性的抗炎化合物,包括 2 种 BET 抑制剂(I-BET151、Ro 11-1464)和 2 种 HDAC 抑制剂(SAHA、TSA);其中 BET 抑制剂可显著上调内源性 CEBPD 及报告基因 CEBPD::SEAP 的 mRNA 表达,强效抑制 IL-6 和 CCL2(I-BET151 分别抑制 65 倍、15 倍,Ro 11-1464 均抑制 9 倍),且 Ro 11-1464 对 IL-1β 无显著影响,安全性更优;HDAC 抑制剂虽抑制内源性 CEBPD,但可上调报告基因 CEBPD::SEAP(推测与报告基因仅含 CEBPD 近端启动子、内源性基因含远端调控区域有关),且抑制 IL-6/CCL2 效率更高(SAHA 抑制 110 倍、15 倍以上,TSA 抑制 130 倍、17 倍以上);所有化合物均无显著细胞毒性,不仅为类风湿关节炎、动脉粥样硬化等炎症驱动疾病提供了 4 种新型候选药物,还揭示了 BET/HDAC 介导的 CEBPD 表观遗传调控差异,为后续靶向 CEBPD 的精准抗炎治疗提供了实验基础和机制参考。
常见细胞污染类型如何辨别及预防解决方法:细胞培养中常见的生物污染类型有7种,分别是细菌污染,支原体污染,原虫污染,黑胶虫污染,真菌污染,病毒污染以及非细胞污染,真菌污染来源,一般是来自实验服,并且具有气候性,多雨······
细胞聚团的原因分析及如何避免:培养物中细胞可能聚集的一些原因包括:1.过度消化、2.环境压力、3.组织分解、4.过度生长、5.污染等;如何避免聚团细胞的生成;首先确认当前细胞生长密度及状态,80%左右的生长密度即可进行······
细胞有空泡原因分析及解决方法:出现细胞空泡情况有1.细胞老化2.培养液错误配制;3.细胞消化时操作不当;4.污染等等,如细胞老化,解决方法,原代细胞使用较低代次进行实验,传代细胞避免传代次数过高···
细胞半换液和全换液操作步骤:第一种方式:细胞全换液;如果是贴壁细胞,可以用全量换液法,直接吸去全部旧培养基,补充足量新鲜完全培养基;第二种方式:细胞半换液;"细胞半换液"又称"细胞半量换液",即弃掉一半旧的培养基,再······
细胞生长缓慢的可能原因有哪些:细胞培养外部因素包括细胞培养基的配方和质量问题,培养条件不理想,污染问题,细胞自身因素包含细胞的健康状态,细胞密度过高或过低,细胞老化现象,细胞特性,当细胞生长出现缓慢的问题时,我······
常用胰腺癌细胞株动物模型及胰腺癌细胞株有哪些:胰腺癌研究中常用的动物模型主要包括化学物质诱导胰腺癌动物模型,基因工程胰腺癌小鼠模型和胰腺癌移植模型,常用的胰腺细胞株MIA-PACA-2人胰腺癌细胞,Capan-2人胰腺癌细······
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
上一篇:蛋白激酶A活性在自发跳动的人源多能干细胞衍生心脏类器官中的时空分析
下一篇:亚洲人专属保护基因!NTCP S267F 变异可降低乙肝进展至肝硬化、肝癌风险
厦门爱恪信生物科技有限公司
手机:15859239971
邮箱:2205839769@qq.com
地址:厦门翔安火炬高新区翔星路96号建业楼D座602

微信公众号
ATCC细胞培养

技术支持
15859239971
Copyright©厦门爱恪信 闽ICP备19027235号-7
公安备案: XML地图
XML地图




 客服QQ
客服QQ