常见细胞污染类型如何辨别及预防解决方法
常见细胞污染类型如何辨别及预防解决方法:细胞培养中常见的生物污染类型有7种,分别是细菌污染,支原体污染,原虫污染,黑胶虫污染,真菌污染,病毒污染以及非细胞污染,真菌污染来源,一般是来自实验服,并且具有气候性,多雨······

发布时间:2025-10-06 08:59:26 细胞资源库平台 访问量:226
荧光素酶报告基因系统是一种基于荧光素酶催化底物氧化反应产生生物发光的检测技术,广泛应用于细胞生物学研究。其中,萤火虫荧光素酶(firefly luciferase, Fluc)因其高灵敏度、宽线性检测范围(约7~8个数量级)以及较短的半衰期(在哺乳动物细胞中约为3小时,在植物细胞中约为3.5小时)而成为最常用的报告基因。其发光信号强度在酶浓度为10⁻¹⁶ mol/L至10⁻⁸ mol/L的范围内与酶活性呈线性关系,并且在理想条件下可检测到低至10⁻²⁰ mol/L的荧光素酶活性。此外,荧光素酶报告基因系统具有非放射性、检测快速、灵敏度高(比氯霉素乙酰转移酶CAT高100倍)等优点,特别适用于高通量筛选和活细胞检测。通过将荧光素酶报告基因载体转染至宿主细胞后,可利用荧光素酶检测系统灵敏且便捷地监测基因表达水平,已成为细胞生物学研究中的重要工具。
英文标题:An all-in-one pipeline for the in vitro discovery and in vivo testing of Plasmodium falciparum malaria transmission blocking drugs
中文标题:疟原虫传播阻断药物的体外发现与体内验证一体化研究平台构建
发表期刊:《Nature Communications》
影响因子:15.7
作者单位:
1. Department of Medical Parasitology and Infection Biology, Swiss Tropical and Public Health Institute, 4123 Allschwil, Switzerland
2. University of Basel, 4001 Basel, Switzerland
3. Department of Medical Microbiology, Radboudumc Center for Infectious Diseases, Radboud University Medical Center, 6525GA Nijmegen, The Netherlands
4. Biology of Host-Parasite Interactions, Department of Parasites and Insect Vectors, Institut Pasteur, CNRS EMR9195, INSERM U1201, Université de Paris-Cité, Paris 75015, France
5. Biomolecular Screening Facility, École Polytechnique Fédérale de Lausanne (EPFL), 1015 Lausanne, Switzerland
6. Epigenetic Chemical Biology, Department of Structural Biology and Chemistry, Institut Pasteur, CNRS UMR3523 Chem4Life, Université de Paris-Cité, Paris 75015, France
7. Medicines for Malaria Venture, 1215 Geneva, Switzerland
8. Present address: Laboratoire de Chimie de Coordination du CNRS UPR8241, LCC-CNRS, Inserm ERL 1289, Université de Toulouse, 31077 Toulouse, France
9. Present address: Global Antibiotic Research and Development Partnership (GARDP), 1202 Geneva, Switzerland
10. Present address: Department of Immunology and Infectious Diseases, Harvard T. H. Chan School of Public Health, Boston, MA 02115, USA
作者信息:
Nicolas M. B. Brancucci¹,², Christin Gumpp¹,², Geert-Jan van Gemert³, Xiao Yu¹,², Armin Passecker¹,², Flore Nardella⁴,⁸, Basil T. Thommen¹,²,¹⁰, Marc Chambon⁵, Gerardo Turcatti⁵, Ludovic Halby⁶, Benjamin Blasco⁷,⁹, Maëlle Duffey⁷,⁹, Paola B. Arimondo⁶, Teun Bousema³, Artur Scherf⁴,⁷, Didier Leroy⁷, Taco W. A. Kooij³, Matthias Rottmann¹,², Till S. Voss¹,²*
疟疾消除需针对疟原虫成熟配子体(仅 V 期可感染蚊子)的传播阻断药物,但现有药物存在局限:伯氨喹(唯一获批药物)对 G6PD 缺乏者有溶血风险,青蒿素类复方(ACTs)对 V 期配子体效果差,且缺乏高效的体外筛选模型(配子体培养同步性差、 viability 检测难)和体内验证模型(无合适动物模拟人疟原虫感染)。本研究通过 CRISPR/Cas9 构建转基因疟原虫株 NF54/iGP1_RE9Hulg8(缺失条件性表达 GDV1 以诱导同步配子体,且 V 期配子体表达红移荧光素酶 RE9H),建立 “体外筛选 - 体内验证 - 蚊子传播阻断” 一体化平台:体外通过发光检测快速筛选 V 期配子体杀伤药物,体内利用人源化 NODscidIL2Ryⁿᵘˡˡ(NSG)小鼠(移植人红细胞)评估药物清除配子体及阻断传播的效果,解决现有研究的技术瓶颈。
通过 CRISPR/Cas9 技术改造 NF54 疟原虫:在 cg6 位点插入条件性 GDV1 过表达 cassette(iGP1)以诱导同步配子体形成,在 ulg8 位点(配子体特异性启动子)插入 2A-RE9H 序列(红移荧光素酶),构建 NF54/iGP1_RE9Hulg8 株;体外培养时,用−GlcN/+Shield-1 诱导配子体分化,加 GlcNAc 清除无性体,获得同步 V 期配子体,建立 96 孔板发光检测 assay(加 D - 荧光素测 RE9H 活性,反映配子体 viability),筛选 4 个化合物库(表观遗传库、激酶抑制剂库、Prestwick 库等共 1740 个分子);
建立人源化 NSG 小鼠模型:每日静脉注射人 AB + 红细胞 11 天,感染 2×10⁸个 V 期配子体,通过尾血涂片显微镜计数和活体成像(IVIS Lumina II)监测配子体循环;体内药物测试中,小鼠感染后 1 天口服给药(伯氨喹、氯喹及临床候选药如 KAE609、KAF156 等),监测 16 天内配子体清除;
通过蚊子饲喂实验(MFA):收集给药后小鼠血液饲喂斯氏按蚊,7 天后解剖计数蚊胃卵囊,验证传播阻断效果;统计用 GraphPad Prism(t 检验 / ANOVA,p<0.05 为显著)。
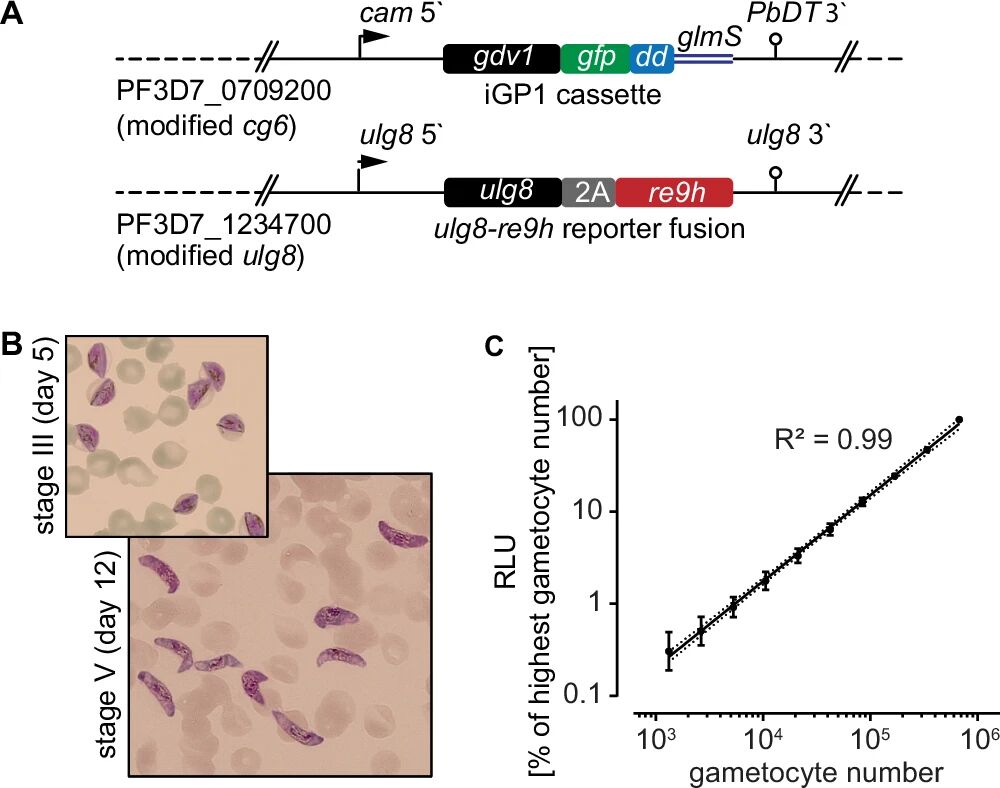
图1:NF54/iGP1_RE9Hulg8 配子体 reporter 株的构建与验证
A 图展示基因组改造位点:cg6 位点插入 iGP1 cassette(调控 GDV1 表达),ulg8 位点插入 2A-RE9H(配子体特异性表达荧光素酶);B 图显微镜观察显示,该菌株可分化为同步的 III 期(第 5 天)和 V 期(第 12 天)配子体,形态正常;C 图显示 V 期配子体数量与 RE9H 发光强度呈强线性相关(R²=0.99),且发光信号随配子体密度增加而升高,证实 RE9H 可精准量化活配子体数量;此外,该菌株无性体增殖率与野生型相当(倍增率 7.0±0.3 vs 6.9±0.8),配子体雌雄比例正常,雄配子体出丝率与野生型无差异,说明改造不影响疟原虫生理功能。
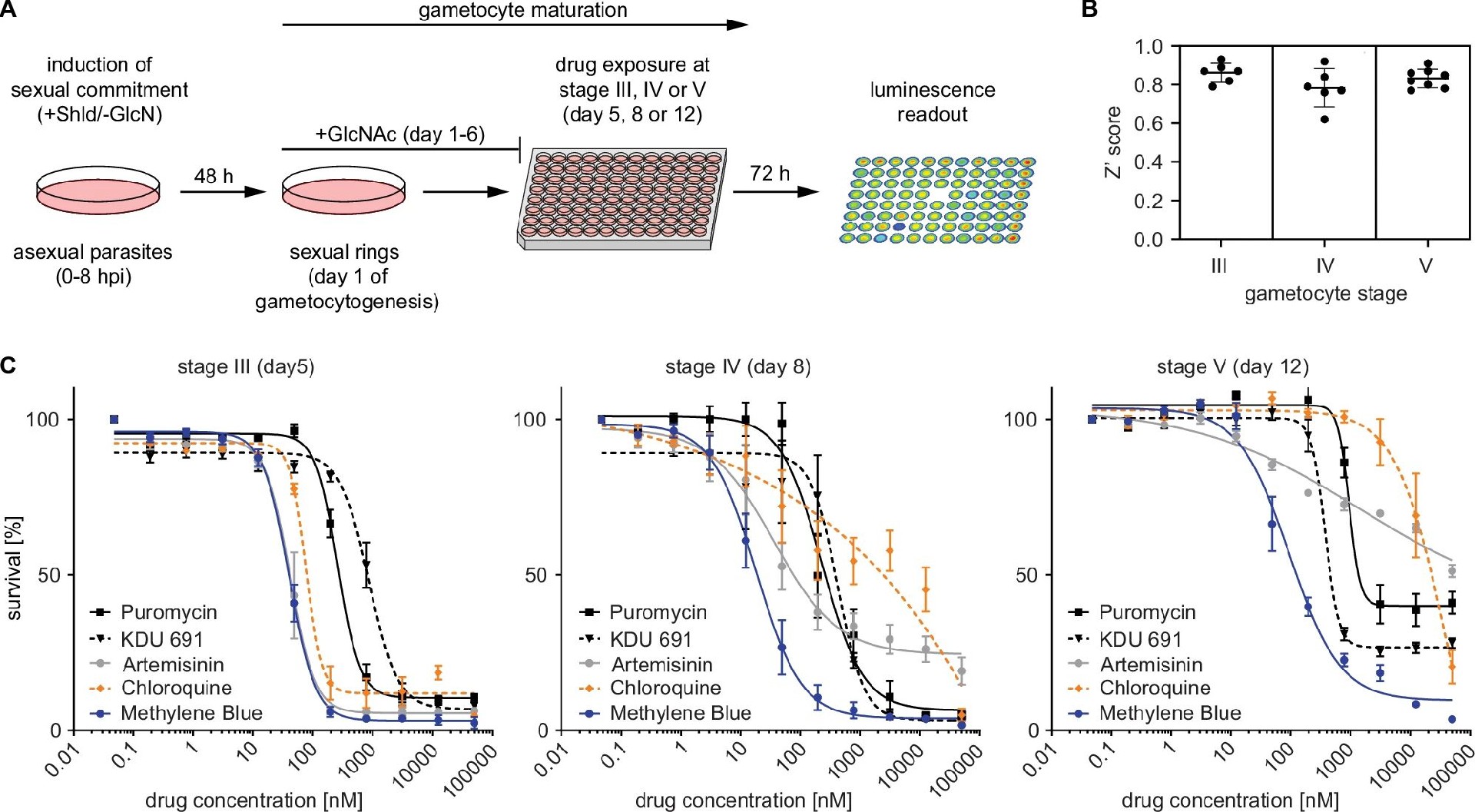
图2:体外配子体杀伤药物筛选 assay 的建立与验证
A 图展示 assay 流程:诱导同步配子体后,在 III 期(第 5 天)、IV 期(第 8 天)、V 期(第 12 天)加药,72 小时后测发光信号;B 图显示 assay 稳定性:Z’值达 0.83±0.04(Z’>0.8 为优秀),说明重复性好;C 图药物验证显示,氯喹仅杀伤 III 期配子体,青蒿素杀伤 III/IV 期但对 V 期无效,而 PI4K 抑制剂 KDU691、亚甲蓝(MB)、嘌呤霉素对所有期次配子体均有杀伤(IC50 见表 S1),与已知药物活性一致,证实 assay 可靠。
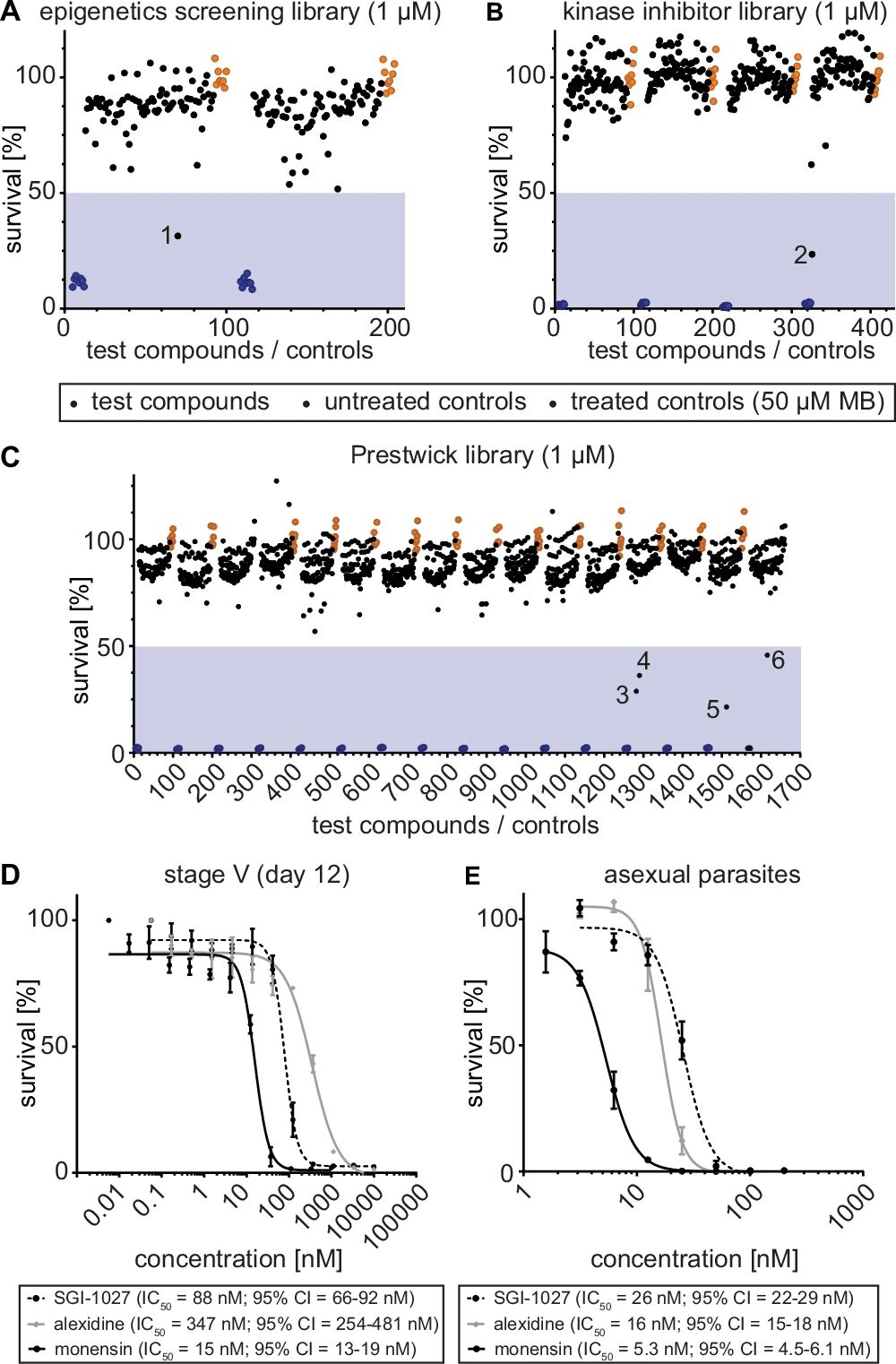
图3:化合物库筛选鉴定 V 期配子体杀伤分子
A-C 图分别为表观遗传库(148 个)、激酶抑制剂库(275 个)、Prestwick 库(1280 个)的筛选结果(1μM 浓度):仅 SGI-1027(表观遗传库,DNMT 抑制剂)、SU4312(激酶抑制剂库,VEGFR2 抑制剂)、莫能菌素、阿来西定、吲哚洛芬、马烯雌酮(Prestwick 库)对 V 期配子体抑制率 > 50%;
D 图剂量反应实验显示,SGI-1027(IC50=88nM)、莫能菌素(IC50=15nM)、阿来西定(IC50=347nM)对 V 期配子体均有纳米级活性;
E 图显示这三种分子同时抑制无性体(IC50 分别为 26nM、5.3nM、16nM),为双活性分子,提示其靶向无性体与配子体共有的通路。
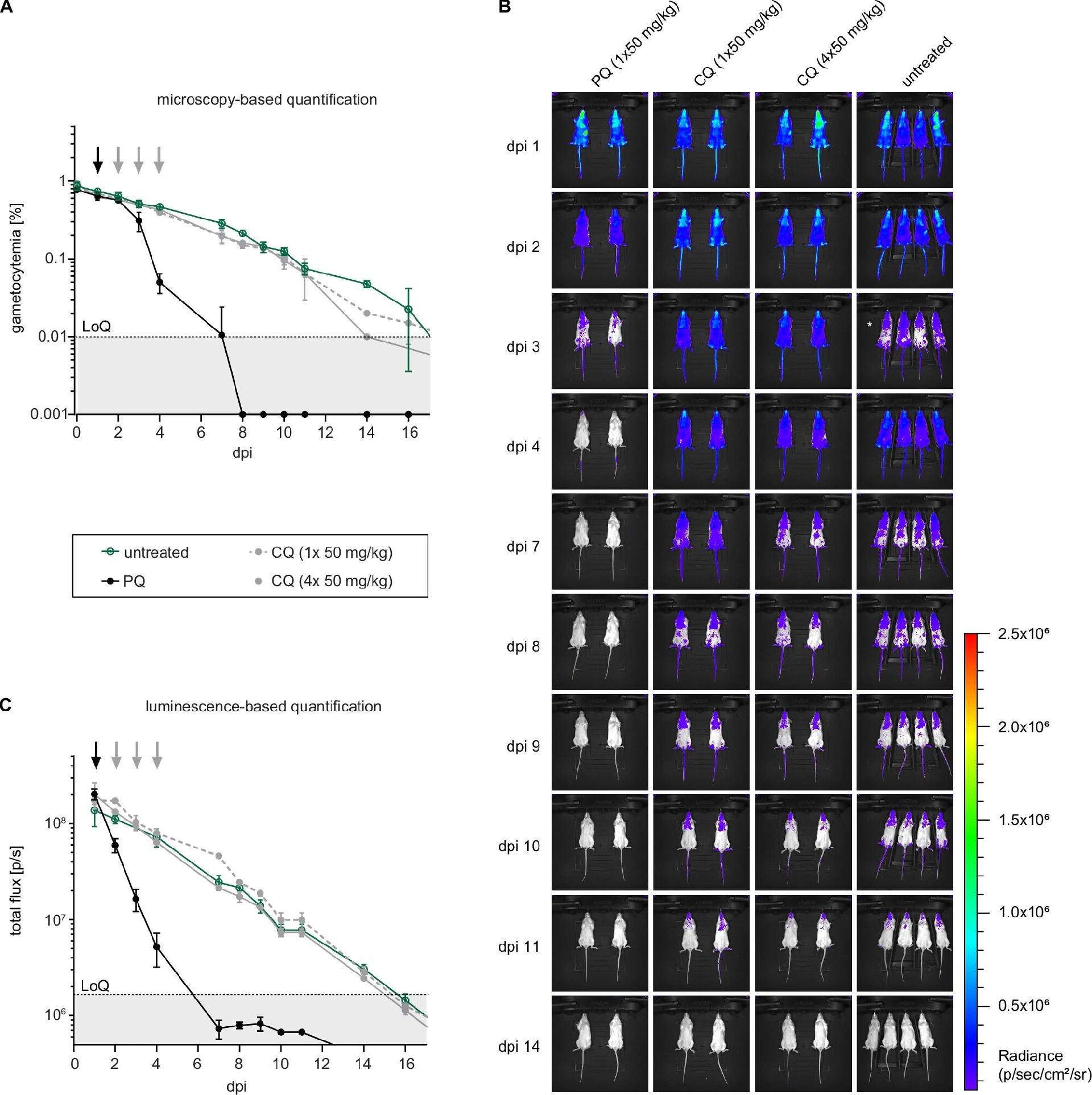
图4:NSG 小鼠模型中配子体的循环与药物清除
A 图显微镜计数显示,未给药小鼠 V 期配子体可在血液中循环 14 天以上(初始配子体血症 0.9%),单剂量氯喹(50mg/kg)或 4 次氯喹给药均无法清除配子体,而单剂量伯氨喹(50mg/kg)在第 7 天将配子体降至检测限以下;
B 图活体成像显示,伯氨喹组发光信号在给药后 1 天即显著下降,第 7 天低于检测限,而氯喹组与未给药组信号无差异;
C 图定量发光通量显示,伯氨喹组总光子通量从 2.5×10⁶ p/s 降至 5×10⁵ p/s 以下,证实该模型可区分药物对 V 期配子体的活性,且发光检测比显微镜更灵敏。
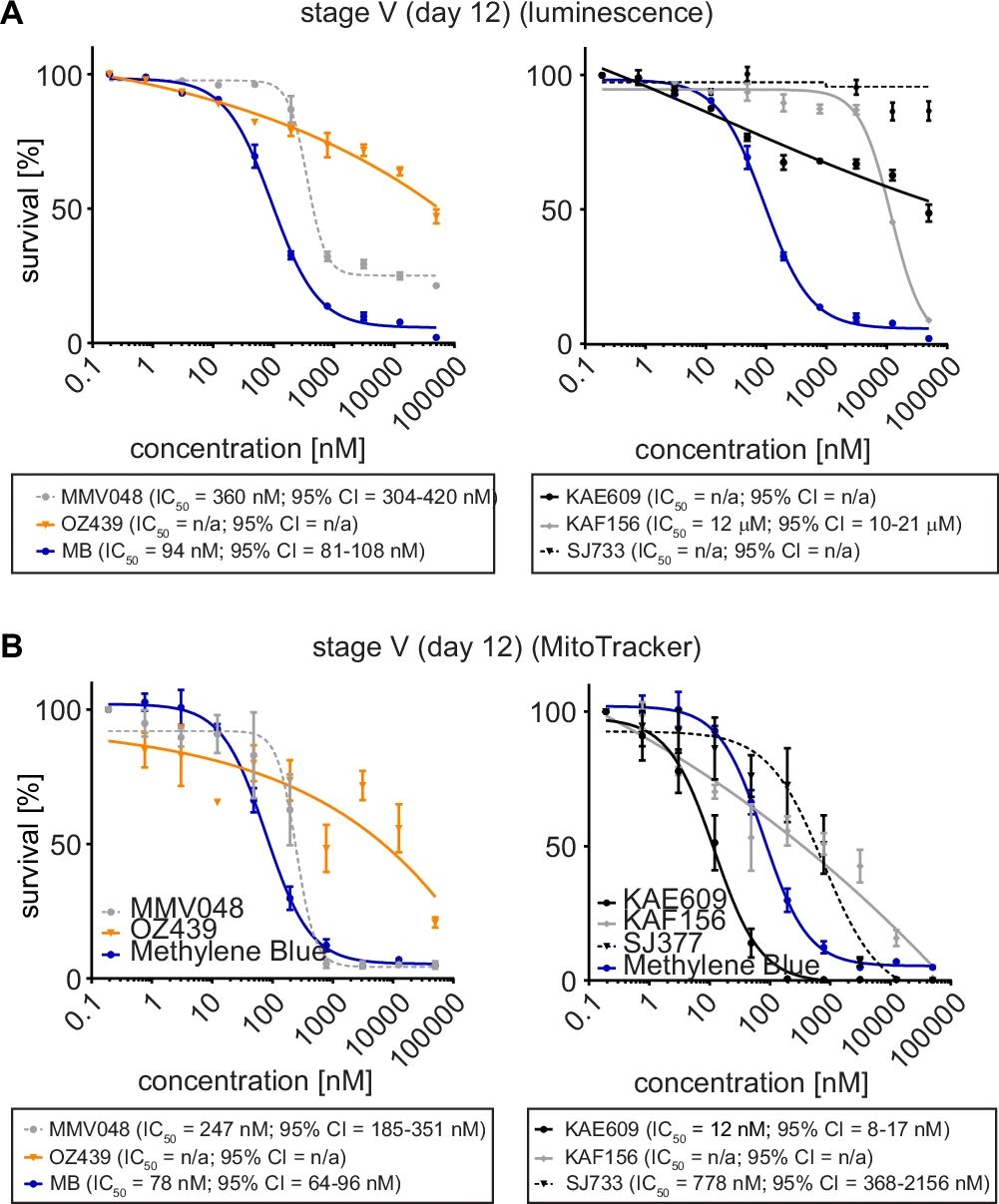
图5:临床候选药物对 V 期配子体的体外活性
A 图发光检测显示,MMV390048(PI4K 抑制剂,IC50=360nM)、亚甲蓝(IC50=94nM)对 V 期配子体有效,而 OZ439(青蒿素衍生物)、KAE609(PfATP4 抑制剂)、KAF156(蛋白分泌抑制剂)、SJ733(PfATP4 抑制剂)活性弱或无活性;
B 图线粒体活性 / 形态检测(MitoTracker 染色)显示,KAE609 实际对 V 期配子体有强活性(IC50=12nM),SJ733 有弱活性(IC50=778nM),原因是 KAE609 虽不影响 RE9H 活性,但会导致配子体形态异常(肿胀、圆形),而发光检测无法反映形态损伤,提示需结合多维度检测;MMV390048、亚甲蓝的两种检测结果一致(IC50 分别为 247nM、78nM),OZ439 仍无活性。
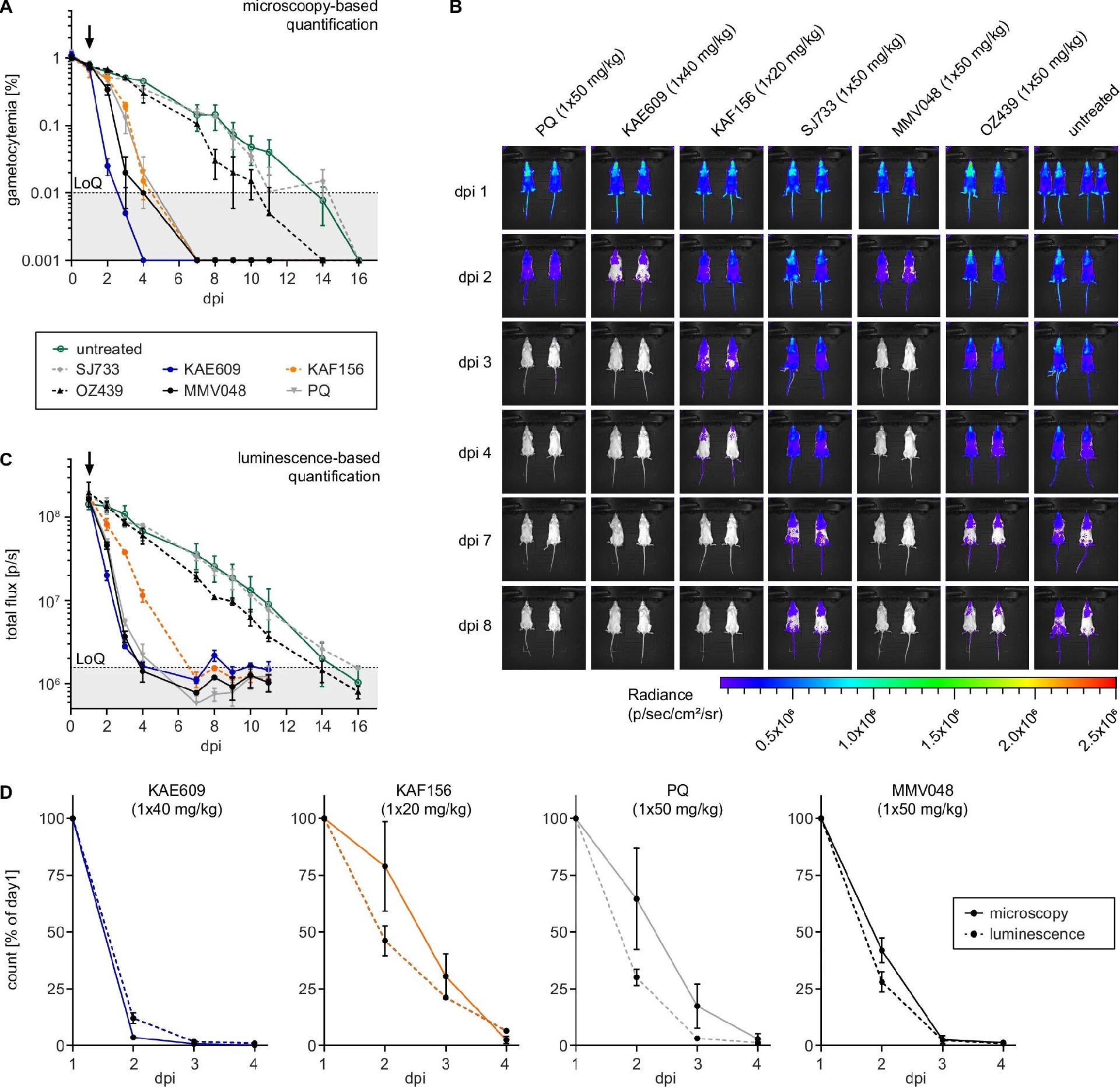
图6:临床候选药物在 NSG 模型中的体内活性
A 图显微镜计数显示,KAF156(20mg/kg)、MMV390048(50mg/kg)、KAE609(40mg/kg)、伯氨喹(50mg/kg)均可清除 V 期配子体:KAE609 在 2 天内将配子体降至检测限以下,KAF156、MMV390048、伯氨喹在 4-7 天内清除,而 SJ733(50mg/kg)、OZ439(50mg/kg)与未给药组无差异;
B 图活体成像显示,KAE609 组发光信号在给药后 1 天即集中于脾脏(腹部左上象限),提示脾脏清除异常配子体;
C 图定量发光通量显示,KAE609 组总光子通量 2 天内下降 95%,MMV390048 组 3 天内下降至检测限,KAF156 组 4 天内下降 80%;
D 图对比显示,发光信号下降速度快于显微镜计数的配子体密度下降,提示药物先使配子体失活(发光降低),后被清除(密度下降)。
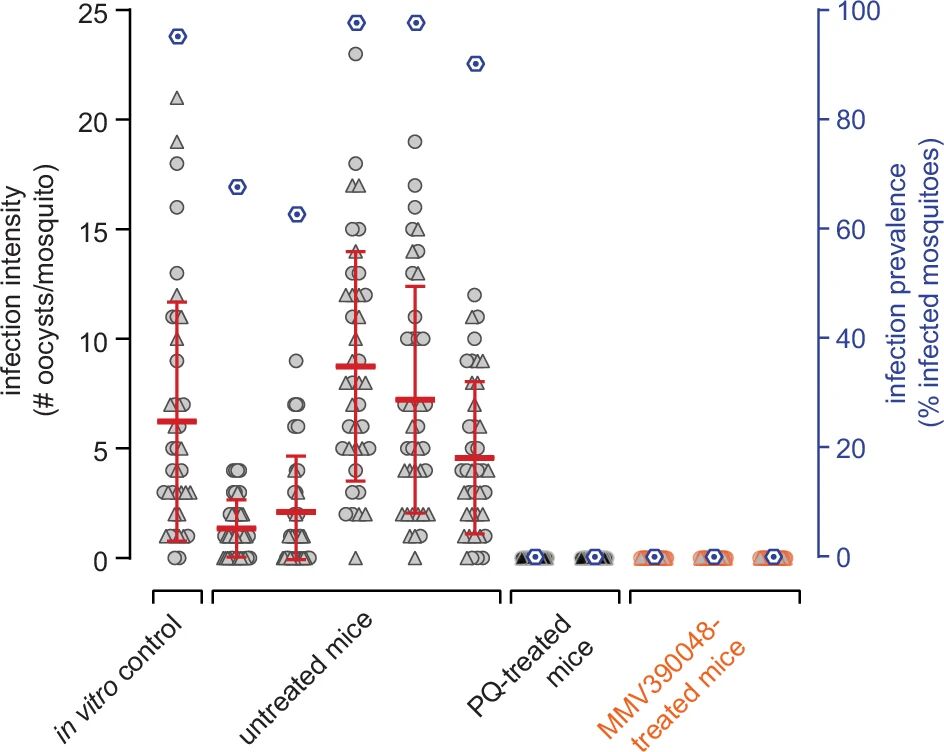
图7:NSG 模型中药物的传播阻断效果验证
通过 MFA 实验,未给药小鼠血液饲喂按蚊后,感染率高(>80%),蚊胃平均卵囊数 > 10 个;而伯氨喹(50mg/kg)、MMV390048(50mg/kg)给药组小鼠血液饲喂的蚊子均无卵囊(感染率 0%),与体外 SMFA(用同一批配子体)结果一致,证实该模型可有效验证药物的体内传播阻断活性,且 MMV390048 与伯氨喹效果相当。
本研究建立的 “IVIS 成像 + Alu PCR” 联合方法,可高效追踪眼部 adMSCs 的短期动态,其中 IVIS 实现非侵入实时监测,Alu PCR 通过人 DNA 检测验证细胞清除,二者互补提升结果可靠性,为眼部细胞治疗的示踪提供标准化技术路径;结膜下注射的 adMSCs 在 7 天内完全清除,无远处器官迁移,且未检测到长期滞留,结合 adMSCs 本身低致瘤性的特点,证实其在眼部应用的短期安全性,为角膜衰竭、眼表炎症等疾病的细胞治疗提供安全依据;但研究存在局限,IVIS 检测限(2×10³ 细胞)可能遗漏少量残留细胞,未来可采用更灵敏的 Akaluc 报告基因,且本研究仅评估 7 天内动态,需进一步探索长期疗效与细胞命运,不同注射剂量对信号稳定性的影响也需后续优化,以完善临床应用方案。
常见细胞污染类型如何辨别及预防解决方法:细胞培养中常见的生物污染类型有7种,分别是细菌污染,支原体污染,原虫污染,黑胶虫污染,真菌污染,病毒污染以及非细胞污染,真菌污染来源,一般是来自实验服,并且具有气候性,多雨······
细胞聚团的原因分析及如何避免:培养物中细胞可能聚集的一些原因包括:1.过度消化、2.环境压力、3.组织分解、4.过度生长、5.污染等;如何避免聚团细胞的生成;首先确认当前细胞生长密度及状态,80%左右的生长密度即可进行······
细胞有空泡原因分析及解决方法:出现细胞空泡情况有1.细胞老化2.培养液错误配制;3.细胞消化时操作不当;4.污染等等,如细胞老化,解决方法,原代细胞使用较低代次进行实验,传代细胞避免传代次数过高···
细胞半换液和全换液操作步骤:第一种方式:细胞全换液;如果是贴壁细胞,可以用全量换液法,直接吸去全部旧培养基,补充足量新鲜完全培养基;第二种方式:细胞半换液;"细胞半换液"又称"细胞半量换液",即弃掉一半旧的培养基,再······
细胞生长缓慢的可能原因有哪些:细胞培养外部因素包括细胞培养基的配方和质量问题,培养条件不理想,污染问题,细胞自身因素包含细胞的健康状态,细胞密度过高或过低,细胞老化现象,细胞特性,当细胞生长出现缓慢的问题时,我······
常用胰腺癌细胞株动物模型及胰腺癌细胞株有哪些:胰腺癌研究中常用的动物模型主要包括化学物质诱导胰腺癌动物模型,基因工程胰腺癌小鼠模型和胰腺癌移植模型,常用的胰腺细胞株MIA-PACA-2人胰腺癌细胞,Capan-2人胰腺癌细······
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
上一篇:BRD4抑制剂有效治疗神经内分泌前列腺癌中的肿瘤谱系可塑性
厦门爱恪信生物科技有限公司
手机:15859239971
邮箱:2205839769@qq.com
地址:厦门翔安火炬高新区翔星路96号建业楼D座602

微信公众号
ATCC细胞培养

技术支持
15859239971
Copyright©厦门爱恪信 闽ICP备19027235号-7
公安备案: XML地图
XML地图




 客服QQ
客服QQ