常见细胞污染类型如何辨别及预防解决方法
常见细胞污染类型如何辨别及预防解决方法:细胞培养中常见的生物污染类型有7种,分别是细菌污染,支原体污染,原虫污染,黑胶虫污染,真菌污染,病毒污染以及非细胞污染,真菌污染来源,一般是来自实验服,并且具有气候性,多雨······

发布时间:2025-09-22 09:06:36 细胞资源库平台 访问量:219
在生物医学研究和药物开发领域,生物发光成像技术因其高信噪比而被广泛应用于细胞测定和动物成像研究。然而,传统的荧光素酶种类有限,限制了同时成像多个分子和细胞事件的能力。为了突破这一限制,科学家们开发了一种新型的ATP非依赖性荧光素酶——NanoLuc(NL),它源自深海虾Oplophorus gracilirostris,并经过工程改造以增强蛋白质稳定性。NanoLuc作为一种小型(19 kDa)、高亮度的荧光素酶,其亮度是传统萤火虫或海肾荧光素酶的100倍,并且使用furimazine作为底物产生明亮的辉光型发光。NanoLuc的意义在于其为双报告基因生物发光分子成像提供了新的可能。它不仅可以在活体小鼠的表层和深层组织中成像,而且其生物发光随时间的变化可以用来定量肿瘤生长,甚至在少量血清中也能检测到分泌的NL。此外,NanoLuc与萤火虫荧光素酶的结合使用,为在完整细胞和活体小鼠中定量TGF-β信号传导的两个关键步骤提供了一种新型双荧光素酶成像策略,从而在正常生理、疾病和药物开发中扩展了信号转导的成像能力。NanoLuc的作用不仅体现在其高灵敏度和高稳定性上,它还具有更小的尺寸,这使得在标记细胞和蛋白质时对样本的侵入性更小,有助于保持细胞或组织的天然状态。NanoLuc的快速反应、低背景发光和多样灵活等特点,使其在生物学和医学研究中具有广泛的应用前景。因此,NanoLuc作为一种新的报告基因,不仅增强了我们对生物过程的理解和疾病机理的研究,而且在开发潜在治疗方法和疗法方面发挥了重要作用。
英文标题:A luminescent attenuated SARS-CoV-2 for the identification and validation of drug-resistant mutants
中文标题:一种用于识别和验证耐药突变体的发光减毒 SARS-CoV-2
发表期刊:《Journal of Virology》
影响因子:3.8
作者单位:
Yao Ma¹, Chengjin Ye¹, Ahmed Magdy Khalil¹,², Sara H. Mahmoud¹,³, Elizabeth B. Sobolik⁴, Alexander L. Greninger⁴, Esteban Castro¹, Nathaniel Jackson¹, Mahmoud Bayoumi¹,⁵, Richard K. Plemper⁶, Luis Martinez-Sobrido¹*
作者信息:
1. Texas Biomedical Research Institute, San Antonio, Texas, USA
2. Faculty of Veterinary Medicine, Zagazig University, Zagazig, Egypt
3. Center of Scientific Excellence for Influenza Viruses, National Research Centre, Giza, Egypt
4. Virology Division, Department of Laboratory Medicine and Pathology, University of Washington Medical Center, Seattle, Washington, USA
5. Virology Department, Faculty of Veterinary Medicine, Cairo University, Giza, Egypt
6. Center for Translational Antiviral Research, Institute for Biomedical Sciences, Georgia State University, Atlanta, Georgia, USA
SARS-CoV-2 变异持续威胁公共卫生,疫苗需不断更新,而抗病毒药物因靶向保守蛋白(如主蛋白酶 Mpro)仍具疗效,但奈玛特韦等 FDA 批准药物的耐药突变已在免疫缺陷患者中发现,亟需安全的耐药突变研究工具。野生型 SARS-CoV-2 筛选耐药突变的实验被归为 “增益功能(GoF)研究”,存在生物安全隐患;此前研究证实缺失 ORF3a 和 7b 的重组 SARS-CoV-2(Δ3a7b)在 K18-hACE2 小鼠和仓鼠中完全减毒,且 ORF3a 缺失会破坏病毒组装效率、降低感染性。为实现耐药突变的高效追踪与安全研究,本研究构建了表达纳米荧光素酶(Nluc)的减毒病毒 Δ3a7b-Nluc,既保留减毒特性,又可通过 Nluc 活性快速监测病毒感染,用于筛选和验证 Mpro 抑制剂奈玛特韦的耐药突变。
通过细菌人工染色体(BAC)系统,将 Nluc 报告基因插入 Δ3a7b 病毒(缺失 ORF3a 和 7b)的核衣壳(N)基因上游,构建 Δ3a7b-Nluc 重组病毒,在表达 hACE2 和 TMPRSS2 的 Vero-AT 细胞中拯救病毒;通过空斑实验、生长动力学(MOI=0.01 感染,24-72h 测病毒滴度)、Nluc 活性检测(48h 达峰)及 EC50 测定(PRNT 法对比奈玛特韦 / 瑞德西韦对 Δ3a7b 与 Δ3a7b-Nluc 的抑制效果)验证病毒特性;将 Δ3a7b-Nluc 在递增浓度奈玛特韦中连续传代 10 次,通过免疫荧光、PRNT、OD560 染色及 Nluc assay 确认耐药性,利用下一代测序(NGS)和 Sanger 测序鉴定 NSP5(编码 Mpro)的突变;通过反向遗传学构建含 L50F、R188G 单突变及双突变的 Δ3a7b-Nluc 病毒,测定其空斑形态、生长动力学及对奈玛特韦的 EC50,验证突变的耐药贡献,同时检测对瑞德西韦的敏感性以排除非特异性耐药。
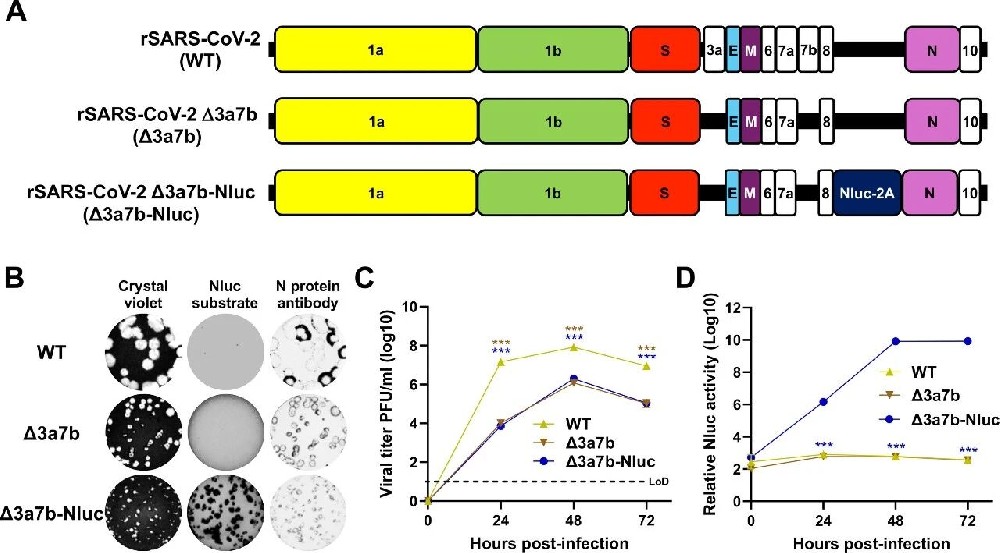
图1:Δ3a7b-Nluc 的构建与表征
A 图 显示病毒基因组结构(Δ3a7b-Nluc 在 Δ3a7b 基础上插入 Nluc-2A 序列);B 图 空斑实验显示,Δ3a7b 与 Δ3a7b-Nluc 空斑均小于野生型(WT),Δ3a7b-Nluc 可通过 Nluc 底物显影;C 图 生长动力学显示,Δ3a7b-Nluc 与 Δ3a7b 复制能力相当,均显著低于 WT;D 图 Nluc 活性与病毒滴度同步,48h 达峰;EC50 测定显示,Δ3a7b-Nluc 对奈玛特韦(0.97μM)和瑞德西韦(5.04μM)的敏感性与 Δ3a7b 一致,与文献中 WT 的 EC50 可比。
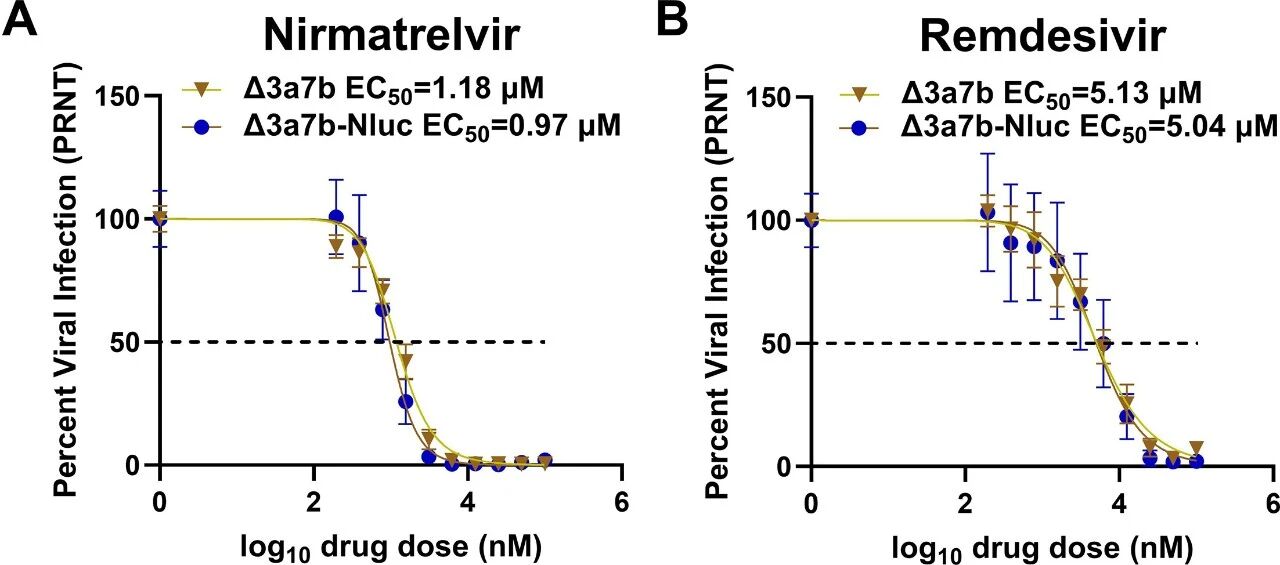
图2:Δ3a7b-Nluc 对 antivirals 的敏感性
PRNT 法测定显示,Δ3a7b-Nluc 对奈玛特韦的 EC50 为 0.97μM,与 Δ3a7b(1.18μM)无显著差异;对瑞德西韦的 EC50 为 5.04μM,同样与 Δ3a7b(5.13μM)可比,且二者 EC50 均与文献中 WT SARS-CoV-2 的数值一致,证实 Δ3a7b-Nluc 可替代 WT 用于 antivirals 敏感性检测。
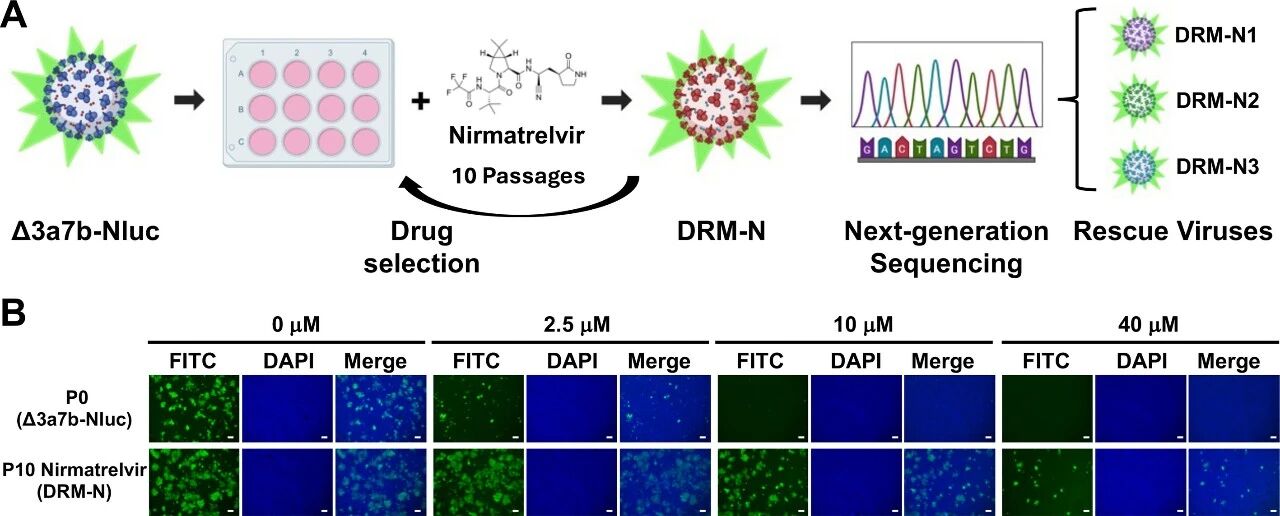
图3:奈玛特韦耐药突变体(DRM-N)的筛选
A 图 展示 Δ3a7b-Nluc 耐药株筛选流程:将 Δ3a7b-Nluc 在含递增浓度奈玛特韦的培养基中连续传代 10 次(P0-P10),仅收集 “药物浓度最高且细胞病变效应(CPE)>50%” 的细胞上清进行传代;P10 代病毒(DRM-N)经 RNA 提取后,通过下一代测序(NGS)和 Sanger 测序鉴定突变,再利用细菌人工染色体(BAC)反向遗传学技术拯救含目标突变的重组病毒。
B 图 为免疫荧光验证结果:Vero-AT 细胞以 MOI=0.001 分别感染 P0 代(亲本 Δ3a7b-Nluc)和 P10 代(DRM-N),加入 0、2.5、10、40μM 奈玛特韦培养 48h 后,用抗 SARS-CoV N 蛋白抗体(1C7C7)和 FITC 标记二抗检测病毒复制,DAPI 染色细胞核(比例尺 = 200μm);结果显示,0μM 药物时两组均有大量 N 蛋白信号(绿色),2.5μM 奈玛特韦可部分抑制 P0 代复制但对 P10 代无影响,10μM 和 40μM 奈玛特韦可完全抑制 P0 代但仅部分抑制 P10 代,证实 P10 代获得奈玛特韦耐药性。
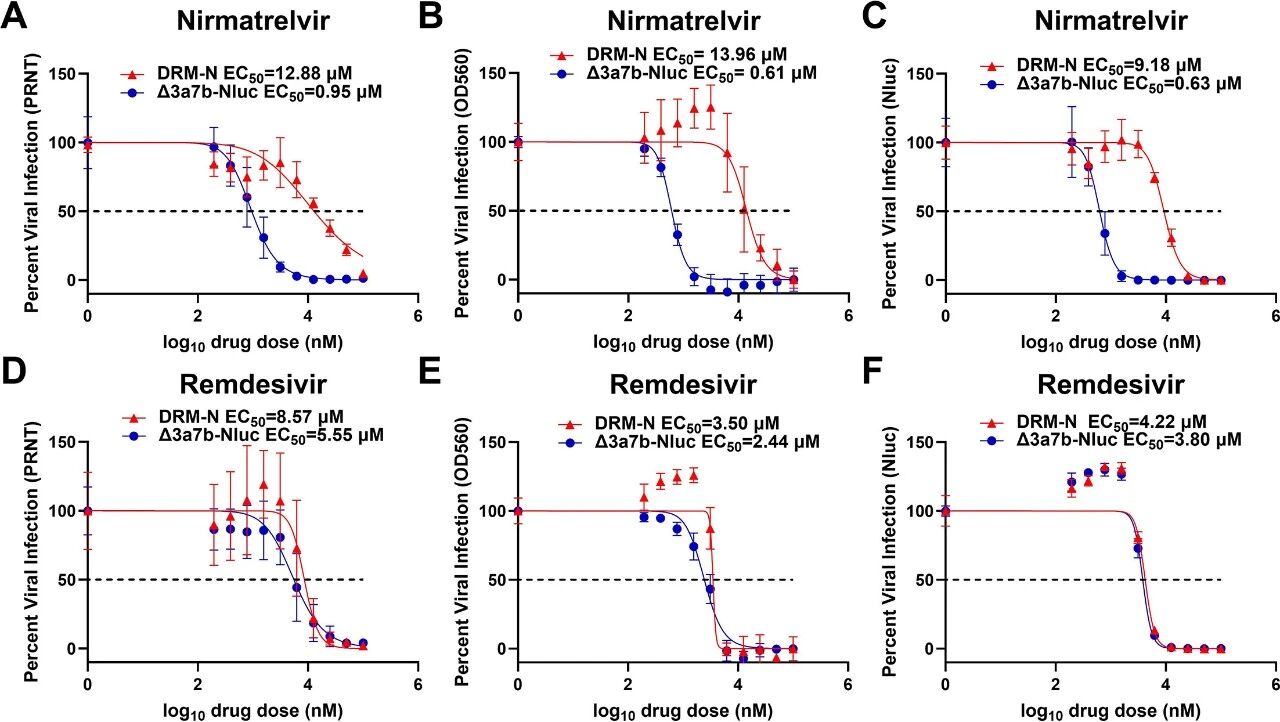
图4:P10 DRM-N 对奈玛特韦的耐药性量化及瑞德西韦敏感性验证
该图通过三种方法量化 DRM-N 对奈玛特韦的耐药性,并验证其对瑞德西韦的敏感性以排除非特异性耐药。
A-C 图 为奈玛特韦耐药量化:PRNT 法(A 图)显示 P0 代 EC50=0.95μM,P10 代 EC50=12.88μM,耐药性提升约 14 倍;OD560 结晶紫染色法(B 图)显示 P0 代 EC50=0.61μM,P10 代 EC50=13.96μM,耐药性提升约 23 倍;Nluc 活性法(C 图)显示 P0 代 EC50=0.63μM,P10 代 EC50=9.18μM,耐药性提升约 15 倍,三种方法结果一致且 Nluc 法操作更高效。
D-F 图 为瑞德西韦敏感性验证:PRNT 法(D 图)、OD560 法(E 图)、Nluc 法(F 图)均显示,P0 代与 P10 代对瑞德西韦的 EC50 无显著差异(如 PRNT 法 P0 代 EC50=0.89μM,P10 代 EC50=0.92μM),证实 DRM-N 的耐药性具有奈玛特韦特异性,不影响对聚合酶抑制剂的敏感性。
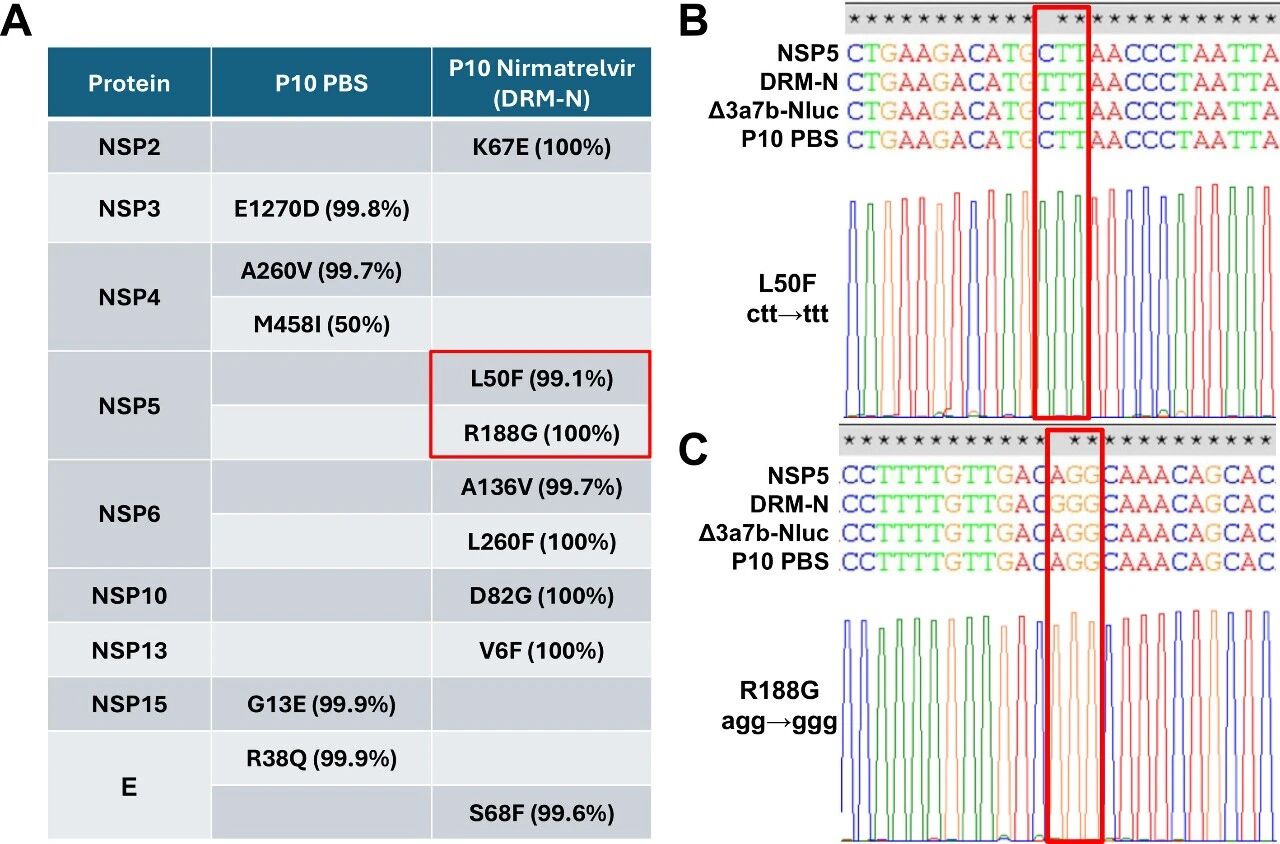
图5:耐药突变的鉴定
NGS 分析显示,P10 DRM-N 在 NSP5 基因中存在 L50F(99.1%)和 R188G(100%)突变,P10 PBS 对照组无此突变;Sanger 测序验证了这两个非同义突变,其他突变(如 NSP2 K67E)因不靶向 Mpro 被排除。
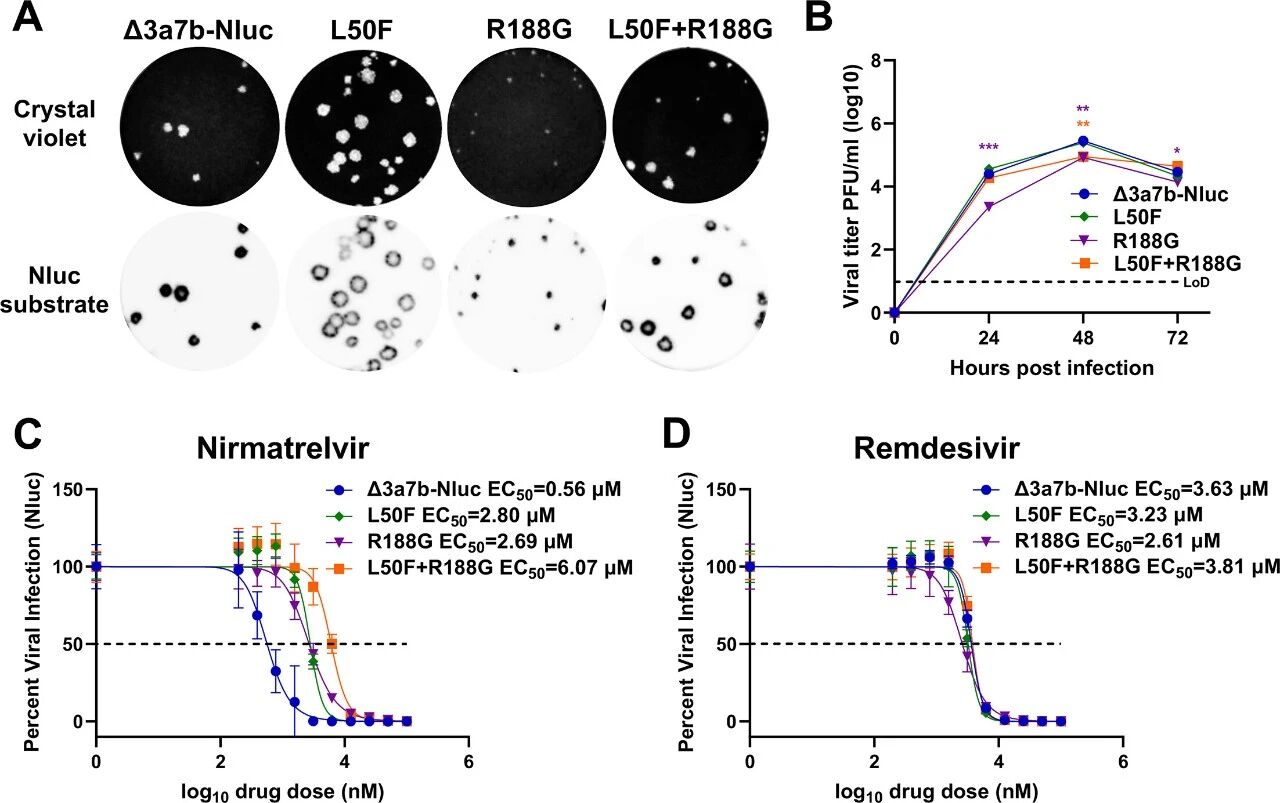
图6:突变的耐药功能验证
构建单突变(L50F、R188G)和双突变(L50F+R188G)病毒:空斑实验显示,R188G 空斑更小,L50F 空斑略大,双突变空斑与亲本一致;生长动力学显示,R188G 复制延迟,双突变呈中间表型;Nluc assay 显示,单突变对奈玛特韦的 EC50 约 2.7μM(5 倍升高),双突变达 6.07μM(11 倍升高),所有突变株对瑞德西韦的 EC50 与亲本无差异,证实 L50F 和 R188G 协同增强奈玛特韦耐药。
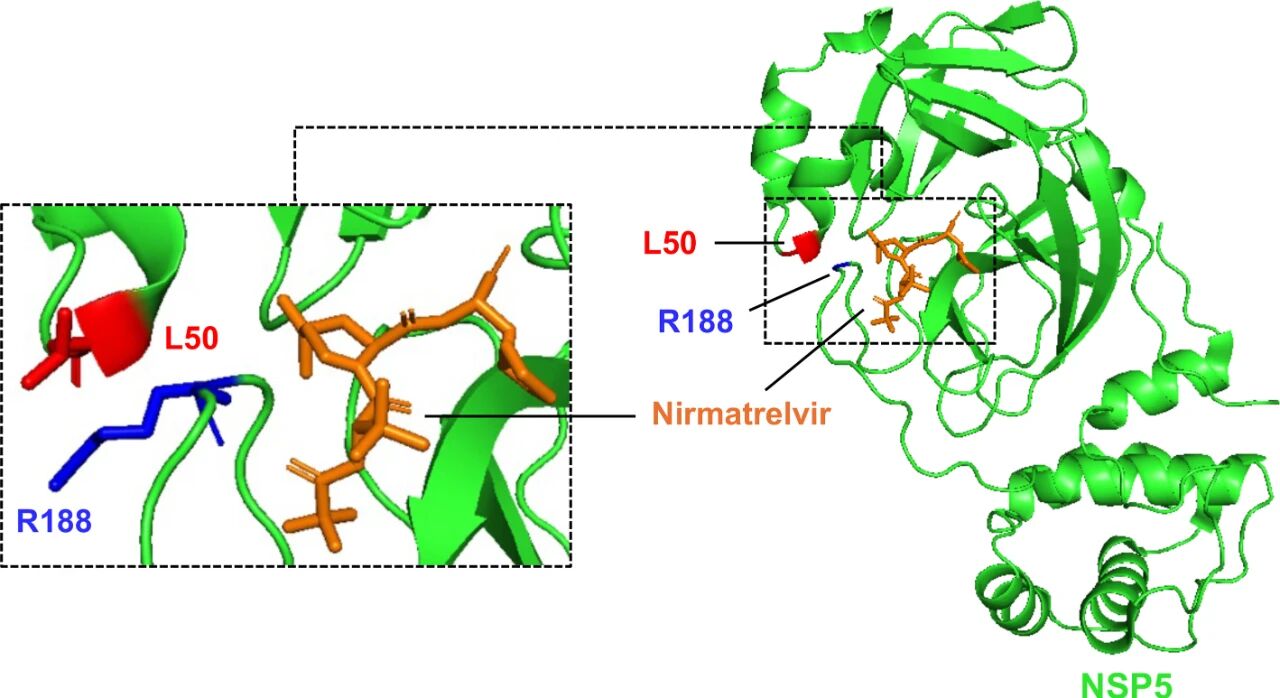
图7:Mpro 与奈玛特韦结合的结构示意图
该图为 SARS-CoV-2 主蛋白酶(Mpro)与奈玛特韦结合的三维结构示意图(基于 PDB ID:7VH8 构建),其中 Mpro 蛋白以绿色丝带模型展示,奈玛特韦以橙色分子结构展示,与耐药相关的两个氨基酸残基 ——L50(位于 Mpro 的疏水 S2 口袋)以红色标记,R188(同样位于 S2 口袋)以蓝色标记。结构分析显示,L50 和 R188 均不直接与奈玛特韦发生相互作用,而是通过维持 S2 口袋的空间构象影响 Mpro 的酶活性:L50F 突变通过引入 bulky 苯丙氨酸残基增强与 Q189 的疏水作用,使 S2 口袋更紧凑;R188G 突变通过替换亲水精氨酸为疏水甘氨酸,同样改变 S2 口袋构象,二者均通过间接影响 Mpro - 底物结合效率导致奈玛特韦耐药,为耐药机制提供结构层面解释。
本研究成功构建了表达 Nluc 的减毒 SARS-CoV-2 Δ3a7b-Nluc,该病毒保留 Δ3a7b 的减毒特性(复制慢、空斑小,无 GoF 风险),Nluc 活性可快速追踪感染,对奈玛特韦 / 瑞德西韦的敏感性与亲本一致;通过 10 次奈玛特韦递增浓度传代,筛选出耐药株 DRM-N,鉴定并验证了 NSP5 的 L50F 和 R188G 突变 —— 单突变分别使奈玛特韦耐药性提升 5 倍,双突变协同提升 11 倍,且不影响对瑞德西韦的敏感性;Δ3a7b-Nluc 为 SARS-CoV-2 耐药突变研究提供了安全高效的工具,避免野生型病毒的 GoF 风险,还可拓展用于疫苗免疫逃逸研究,为抗病毒药物研发和耐药监测提供支撑。
常见细胞污染类型如何辨别及预防解决方法:细胞培养中常见的生物污染类型有7种,分别是细菌污染,支原体污染,原虫污染,黑胶虫污染,真菌污染,病毒污染以及非细胞污染,真菌污染来源,一般是来自实验服,并且具有气候性,多雨······
细胞聚团的原因分析及如何避免:培养物中细胞可能聚集的一些原因包括:1.过度消化、2.环境压力、3.组织分解、4.过度生长、5.污染等;如何避免聚团细胞的生成;首先确认当前细胞生长密度及状态,80%左右的生长密度即可进行······
细胞有空泡原因分析及解决方法:出现细胞空泡情况有1.细胞老化2.培养液错误配制;3.细胞消化时操作不当;4.污染等等,如细胞老化,解决方法,原代细胞使用较低代次进行实验,传代细胞避免传代次数过高···
细胞半换液和全换液操作步骤:第一种方式:细胞全换液;如果是贴壁细胞,可以用全量换液法,直接吸去全部旧培养基,补充足量新鲜完全培养基;第二种方式:细胞半换液;"细胞半换液"又称"细胞半量换液",即弃掉一半旧的培养基,再······
细胞生长缓慢的可能原因有哪些:细胞培养外部因素包括细胞培养基的配方和质量问题,培养条件不理想,污染问题,细胞自身因素包含细胞的健康状态,细胞密度过高或过低,细胞老化现象,细胞特性,当细胞生长出现缓慢的问题时,我······
常用胰腺癌细胞株动物模型及胰腺癌细胞株有哪些:胰腺癌研究中常用的动物模型主要包括化学物质诱导胰腺癌动物模型,基因工程胰腺癌小鼠模型和胰腺癌移植模型,常用的胰腺细胞株MIA-PACA-2人胰腺癌细胞,Capan-2人胰腺癌细······
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
下一篇:眼部ad MSCs追踪新方案IVIS+Alu PCR联手,7 天摸清细胞去向为眼疗安全保驾护航
厦门爱恪信生物科技有限公司
手机:15859239971
邮箱:2205839769@qq.com
地址:厦门翔安火炬高新区翔星路96号建业楼D座602

微信公众号
ATCC细胞培养

技术支持
15859239971
Copyright©厦门爱恪信 闽ICP备19027235号-7
公安备案: XML地图
XML地图




 客服QQ
客服QQ