常见细胞污染类型如何辨别及预防解决方法
常见细胞污染类型如何辨别及预防解决方法:细胞培养中常见的生物污染类型有7种,分别是细菌污染,支原体污染,原虫污染,黑胶虫污染,真菌污染,病毒污染以及非细胞污染,真菌污染来源,一般是来自实验服,并且具有气候性,多雨······

发布时间:2025-06-20 09:00:00 细胞资源库平台 访问量:284
英文标题:A cocktail nanovaccine targeting key entry glycoproteins elicits high neutralizing antibody levels against EBV infection
中文标题:一种靶向关键进入糖蛋白的鸡尾酒纳米疫苗可诱导高水平中和抗体以抵抗EBV感染
发表期刊:《Nature Communications》
影响因子:14.7
作者单位:中山大学肿瘤防治中心、河南大学、重庆医科大学等
作者信息:
Ling Zhong, Wanlin Zhang, Hong Liu, Xinyu Zhang, Zeyu Yang, Zhenfu Wen, Ling Chen, Haolin Chen, Yanran Luo, Yanhong Chen, Qisheng Feng, Mu-Sheng Zeng, Qiujian Zhao, Lixin Liu, Claude Krummenacher, Yi-Xin Zeng, Yongming Chen, Miao Xu, Xiao Zhang
1. EBV的疾病负担
EBV感染全球超过95%的成年人,与多种恶性肿瘤(如伯基特淋巴瘤、鼻咽癌)和自身免疫性疾病(如多发性硬化症)密切相关。
每年导致约23万-35万新发癌症病例和13万-20万死亡病例,但目前尚无预防性疫苗或有效治疗药物。
2. 疫苗开发的挑战
传统疫苗(如基于gp350的疫苗)在临床试验中未能实现完全保护。
EBV通过复杂的生命周期感染B细胞和上皮细胞,需靶向多种糖蛋白(如gHgL、gB、gp42)以阻断感染。
3. 纳米疫苗的优势
纳米载体可协同递送抗原和佐剂(CpG/MPLA),增强免疫原性并避免全身毒性。
靶向淋巴结运输,促进树突细胞成熟和生发中心反应,诱导持久的体液与细胞免疫。
1.纳米疫苗设计
抗原选择: gHgL(介导上皮细胞感染)、gB(融合蛋白)、gp42(介导B细胞感染)。
载体系统: DOTAP/PLGA/DSPE-PEG2000纳米颗粒共包裹抗原与佐剂,形成核心-壳结构(粒径72-89 nm)。
2.免疫策略
动物模型: C57BL/6J小鼠和新西兰白兔,皮下或肌肉注射三次。
对照组: 游离抗原(Free-form)、铝佐剂疫苗(Alum)、空载体纳米颗粒(eNP)。
3.免疫效果评估
体液免疫: ELISA检测抗原特异性IgG/IgA,中和实验评估B细胞和上皮细胞感染抑制。
细胞免疫: 流式检测记忆B细胞、效应T细胞(IFN-γ/TNF-α/IL-2)及生发中心形成。
保护效果: 人源化小鼠模型(SCID-beige)被动免疫后,检测EBV病毒载量和淋巴瘤发生。
1. 纳米疫苗特性
高包封率(抗原>84%,佐剂近100%),缓释24小时(图1)。
高效靶向淋巴结,促进树突细胞成熟(CD80/CD86/MHC-II表达上调)(图3)。
2. 免疫应答增强
体液免疫: 纳米疫苗诱导的IgG中和效价显著高于游离抗原(B细胞:2.1 vs 1.5 log10;上皮细胞:2.9 vs 2.1 log10)(图2)。
细胞免疫: 效应记忆T细胞(CD44hiCD62Llow)比例增加2倍,生发中心B细胞扩增(图2, 3)。
3. 鸡尾酒疫苗优势
广谱中和: NP-cocktail(gHgL+gB+gp42)的中和效价较单抗原疫苗提高3-5倍(图4)。
持久保护: 人源化小鼠模型中,NP-cocktail IgG完全阻断EBV病毒血症和淋巴瘤(图6, 7)。
4. 机制解析
抗体靶向关键表位(如gHgL的AMMO1表位、gB的3A5表位),与天然感染中高效中和抗体重叠(图4J-L)。
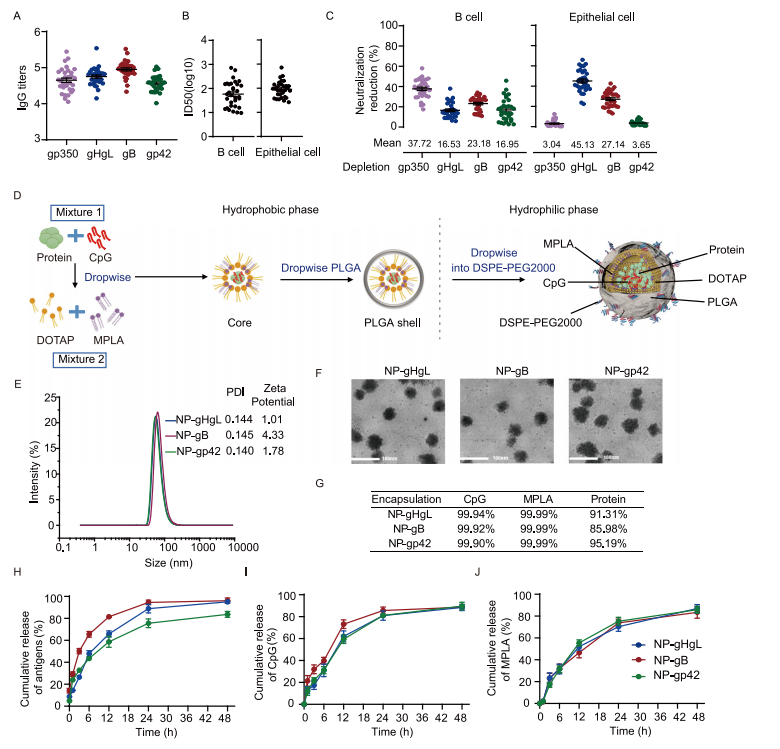
图1:gHgL、gB和gp42是开发针对EBV感染的纳米疫苗的潜在抗原
图(A) 32名健康EBV携带者血清中的抗gp350、抗gHgL、抗gB和抗gp42 IgG滴度。数据点表示为平均值±标准误。
图(B)从32名健康EBV携带者收集的血清对Akata B细胞(左图)和HNE1细胞(右图)的CNE2-EBV感染的中和效果。通过S形趋势拟合计算半最大抑制稀释倍数(ID50)。数据点表示为平均值±标准误。
图(C)通过耗尽糖蛋白特异性抗体后,血清对Akata B细胞和HNE1细胞的EBV感染中和能力的减少。中和能力减少的百分比计算为(1-ID50-耗尽/ID50-前)×100%。数据点表示为平均值±标准误(n = 32)。
图(D)纳米疫苗制备的示意图。图中的元素由Biorender.com制作。E NP-gHgL、NP-gB和NP-gp42纳米疫苗的粒径、多分散指数(PDI)和zeta电位。
F NP-gHgL、NP-gB和NP-gp42纳米疫苗的透射电子显微镜(TEM)图像(比例尺= 100 nm)(n = 3)。
图(G)将抗原和佐剂封装到NP-gHgL、NP-gB和NP-gp42中的效率。每个纳米疫苗的平均封装百分比在三次独立实验中显示。体外抗原(H)、CpG (I)和MPLA (J)的累积释放情况。NP-gHgL、NP-gB和NP-gp42在PBS中与Tween 80混合,并在37°C下以100 rpm的速度连续摇动孵育。数据代表每个纳米疫苗三次独立实验的平均值±标准误。源数据文件中提供了原始数据。DOTAP,1,2-二油酰基-3-三甲基铵丙烷;MPLA,单磷酸脂质A;PLGA,聚(乳酸-共-乙醇酸);DSPE-PEG2000,1,2-二硬脂酰-sn-甘油-3-磷酸乙醇胺-N-[(聚乙二醇)−2000]。
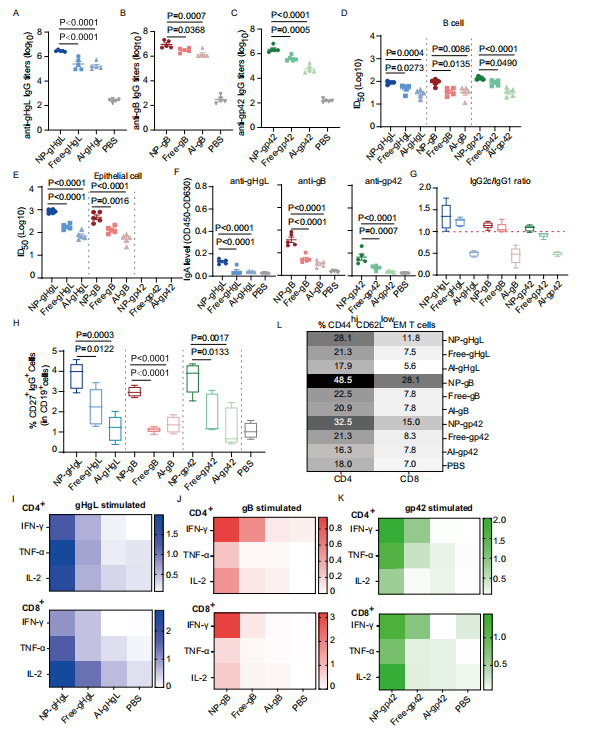
图2:纳米疫苗诱导了强大的体液和细胞免疫反应
图(A) NP-gHgL、Free-gHgL、Al-gHgL和PBS分别诱导的抗gHgL IgG滴度。
图(B) NP-gB、Free-gB、Al-gB和PBS分别诱导的抗gB IgG滴度。
图(C) NP-gp42、Free-gp42、Al-gp42和PBS分别诱导的抗gp42 IgG滴度。
通过使用不同疫苗配方免疫的C57BL/6 J小鼠,在第35天收集的血清能够中和CNE2-EBV感染Akata B细胞(D)和HNE1上皮细胞(E)。半最大抑制稀释度(ID50)是通过S型趋势拟合计算得出的。基于gp42的疫苗诱导的血清无法在体外中和EBV对上皮细胞的感染。
图(F)用不同疫苗配方免疫的C57BL/6 J小鼠在第35天的血清IgA水平。
图(G)用不同疫苗配方免疫的C57BL/6 J小鼠在第35天的血清IgG2c/ IgG1比率(n=5)。H用不同疫苗配方免疫的C57BL/6 J小鼠在第35天脾脏中的记忆B细胞通过流式细胞术评估(n=5)。
图(H)在第35天,通过流式细胞术评估了用不同疫苗配方免疫的C57BL/6 J小鼠脾脏中的记忆B细胞(n = 5)。通过细胞内细胞因子染色测定法,在体外用gHgL (I)、gB (J)和gp42 (K)重新刺激后,测量了第35天脾脏中抗原特异性CD4+(上图)和CD8+(下图)T细胞的反应。数字表示在体外用特定抗原重新刺激后,CD4+ T细胞或CD8+ T细胞中IFN-γ+、TNF-α+或IL-2+细胞的百分比。数据显示为CD4+ T细胞或CD8+ T细胞中IFN-γ+、TNF-α+或IL-2+细胞的平均百分比(n = 5)。第35天,通过流式细胞术评估了脾脏中的效应记忆(EM)CD4+ T细胞(左图)和CD8+ T细胞(右图)(策略见图S12)。数据显示为CD4+和CD8+ EM T细胞的平均百分比(n = 5)。A-F数据点显示为平均值±标准误(n = 5)。G,H中线表示中位数,上、下箱线表示四分位数;须线表示最大和最小值。P值通过单因素方差分析与Dunnett多重比较计算得出,并以精确值显示。源数据作为源数据文件提供。
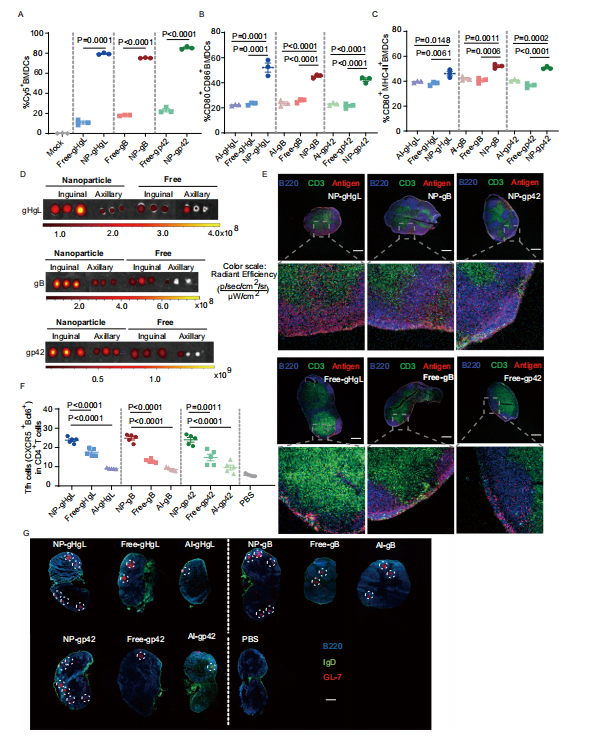
图3:纳米疫苗递送至淋巴结,增强骨髓来源的树突状细胞(BMDCs)的摄取和成熟,以及生发中心(GC)的形成
图(A)Cy5+标记的BMDCs在与PBS(对照组)、游离或纳米颗粒形式的Cy5标记gHgL、gB和gp42孵育4小时后,通过流式细胞术在三个独立实验中测定阳性细胞的比例(n = 3)。检测了与游离(抗原与MPLA和CpG佐剂混合)、铝盐(抗原与无纳米颗粒的铝盐)或纳米颗粒(纳米颗粒与CpG、MPLA和抗原)形式的gHgL、gB和gp42孵育24小时的BMDCs上CD80和CD86 (B)及MHC-II和CD86 (C)的表达情况(n = 3)。
图(D)小鼠接种含有Cy5标记抗原的疫苗后,其淋巴结(LNs)图像(n = 3)。C57BL/6 J小鼠被注射10微克游离或纳米颗粒形式的Cy5标记gHgL、gB或gp42。在6小时后,采集腋窝(远离疫苗注射点)和腹股沟(靠近疫苗注射点)的淋巴结,并使用IVIS光学成像系统测量Cy5荧光(n = 3)。E免疫后6小时腹股沟淋巴结的共聚焦显微镜图像(比例尺= 400微米)。淋巴结(LNs)用Alexa Fluor 594-B220染色以检测B细胞,用FITC-CD3染色以检测T细胞(n = 3)。在第35天,通过不同疫苗配方免疫的小鼠脾脏中生成了F Tfh细胞。每个组包含五只雌性C57BL/6 J小鼠(n = 5)。
图(G)显示了通过不同疫苗配方免疫的C57BL/6 J小鼠腹股沟淋巴结中的生发中心形成情况。在首次三次免疫后的第35天收集淋巴结,并用Alexa Fluor 594-IgD、Alexa Fluor 488-B220和Alexa Fluor 647-GL7染色(比例尺= 400微米)(n=5)。NP-:含有抗原和两种佐剂CpG和MPLA的纳米疫苗。Free-:抗原与两种佐剂CpG和MPLA混合。Al-:抗原与铝佐剂混合。
C和F的数据以平均值±标准误的形式展示。源数据文件中提供了原始数据。P值使用未配对双尾Welch‘s t检验计算,精确值显示。B、C和F的P值使用单因素方差分析(ANOVA)与Dunnett’s多重比较法计算,精确值显示。
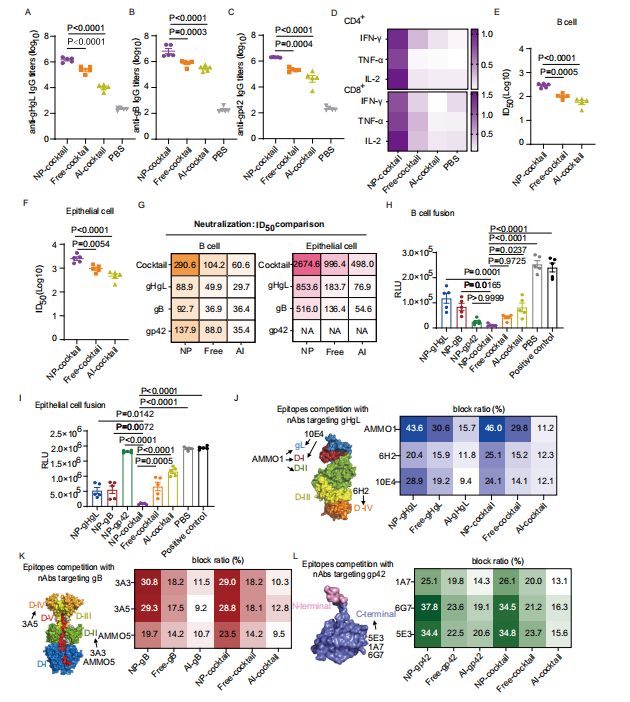
图4:鸡尾酒纳米疫苗在小鼠中对EBV抗原激发了更强烈的免疫反应
图(A)第35天从接种不同疫苗配方的C57BL/6 J小鼠中收集的血清,检测了总抗gHgL(A)、抗gB IgG (B)和抗gp42 IgG (C)的滴度。
图(D)显示了接种指定疫苗的小鼠脾脏中,在第35天(5周)时的抗原特异性CD4+(上图)和CD8+(下图)T细胞反应。通过体外用gHgL、gB和gp42重新刺激后,使用细胞内细胞因子染色测定法测量了抗原特异性T细胞(n = 5)。数字表示体外重新刺激后,CD4+ T细胞或CD8+ T细胞中IFN-γ+、TNF-α+或IL-2+细胞的百分比(n = 5)。
图(E)显示了接种不同疫苗配方的五只C57BL/6 J小鼠在第35天收集的血清对Akata B细胞的CNE2-EBV感染的中和效果。
图(F)显示了接种不同疫苗配方的五只C57BL/6 J小鼠在第35天收集的血清对Akata-EBV感染HNE1上皮细胞的中和效果。
图(G)比较了第35天从接种混合疫苗或单独疫苗的C57BL/6 J小鼠中收集的血清中的B细胞中和滴度(左)和上皮细胞中和滴度(右)。通过S形趋势拟合计算出的半最大抑制稀释倍数(ID50)值。数据显示为平均ID50(n = 5)。不同疫苗配方接种的C57BL/6 J小鼠在第35天收集的血清对B细胞-细胞融合(H)和上皮细胞-细胞融合(I)的抑制效果。未与任何血清孵育的效应细胞作为阳性对照。RLU,相对发光单位。通过ELISA测定了血清抗体与gHgL特异性中和抗体(J)、gB特异性中和抗体(K)和gp42特异性中和抗体(L)的竞争结合能力。糖蛋白结构显示了中和表位的位置。数据代表了每种指定的单克隆中和抗体与第35天从接种不同疫苗配方的C57BL/6 J小鼠中收集的血清抗体竞争的能力。阻断比率显示为每种条件下的平均阻断比率(n = 5)。源数据以源数据文件形式提供。A-C、E、F、H和I数据点显示为平均值±标准误(n = 5)。P值通过单因素方差分析结合Dunnett多重比较计算得出,并以精确值形式展示。
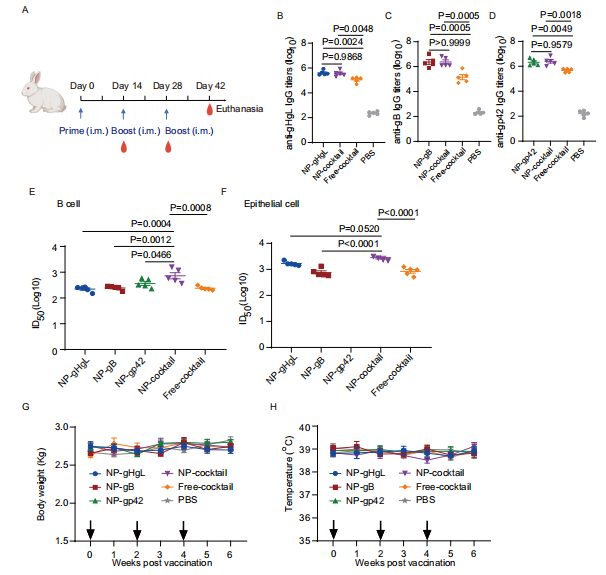
图5:兔子免疫及体液免疫反应的诱导
图(A)兔子免疫时间表。该图基于Biorender.com制作。兔子在第0天、第14天和第28天通过肌肉注射不同疫苗配方三次,剂量见表S2。分别在第0天、第14天、第28天和第42天采集血液样本。兔子在第42天被实施安乐死。B-D用不同疫苗配方免疫的兔子在第42天采集的血清IgG滴度。
图(B)由NP-gHgL、NP-鸡尾酒、Free-鸡尾酒和PBS诱导的抗gHgL IgG滴度(C)由NP-gB、NP-鸡尾酒、Free-鸡尾酒和PBS诱导的抗gB IgG滴度。
图(D)由NP-gp42、NP-鸡尾酒、Free-鸡尾酒和PBS诱导的抗gp42 IgG滴度。
图(E)用不同疫苗配方免疫的兔子在第42天收集的血清对Akata B细胞的CNE2-EBV感染的中和效果。
图(F)用不同疫苗配方免疫的兔子对HNE1上皮细胞的Akata-EBV感染的中和效果。通过S形趋势拟合计算半最大抑制稀释倍数(ID50)值。
图(G)兔子接种指定疫苗配方后的体重(H)和体温(G)被监测。黑色箭头指示免疫时间。B-H数据点显示为平均值±标准误(n = 5)。B-F使用单因素方差分析与Turkey多重比较计算的P值以精确值显示。源数据作为源数据文件提供。
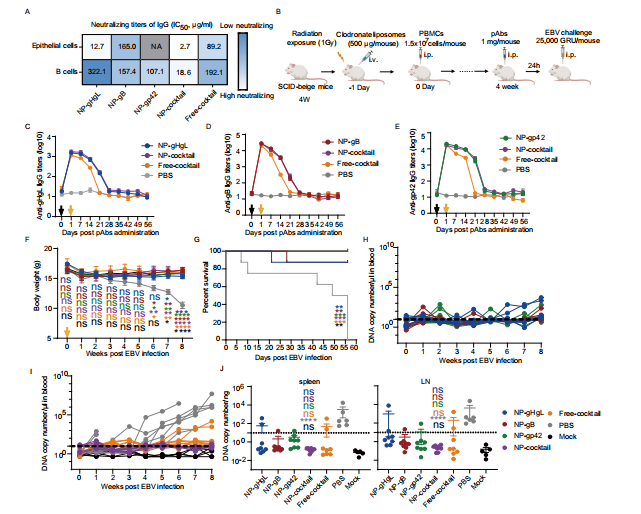
图6:免疫后从兔血清中纯化的总IgG抗体,保护人源化小鼠免受致命EBV挑战
图(A)上图显示了用指定疫苗免疫的三只兔子血清中纯化的IgG对Akata-EBV感染HNE1上皮细胞(顶部)和CNE2-EBV感染Akata B细胞(底部)的中和效果。半最大抑制浓度(半抑制浓度)通过S型趋势拟合计算得出。
图(B)人源化小鼠重建及被动保护对EBV致命挑战的示意图。i.v.表示静脉注射;i.p.表示腹腔注射。实验使用了53只雌性SCID-beige小鼠。该图基于Biorender.com创建。人源化小鼠血清中的抗gHgL(C)、抗gB(D)和抗gp42(E)兔IgG滴度。黑色箭头指示IgG的给药,黄色箭头指示EBV的给药。数据以平均值±标准误(n = 8)表示。F EBV挑战后小鼠的体重。黄色箭头指示EBV感染的日期。数据以平均值±标准误表示。模拟组的人源化小鼠未接受EBV挑战。
图(G)人类化小鼠EBV挑战后的生存曲线。P值见源数据文件。星号的颜色表示统计学差异。
图(H、I)组外周血中的EBV DNA拷贝数。每条线代表一只小鼠,虚线表示检测限。H、I总结了同一实验,为了清晰起见,数据被分为两个图表。J组显示了不同IgG处理的人类化小鼠脾脏(左图)和肠系膜淋巴结(右图)中的EBV DNA拷贝数。虚线表示检测限。由于样本在第7天和第10天死亡,未收集PBS组中两只小鼠的样本,因为样本已腐烂。数据以平均值±标准误的形式展示(NP-gHgL、NP-gB、NP-gp42、NP-鸡尾酒和Free-鸡尾酒组n=8,PBS组n = 6,模拟组n=5)。F、J使用单因素方差分析(ANOVA)和Dunnett多重比较进行了统计分析,精确的P值见源数据文件。星号或ns表示统计学差异。源数据文件中提供了源数据。
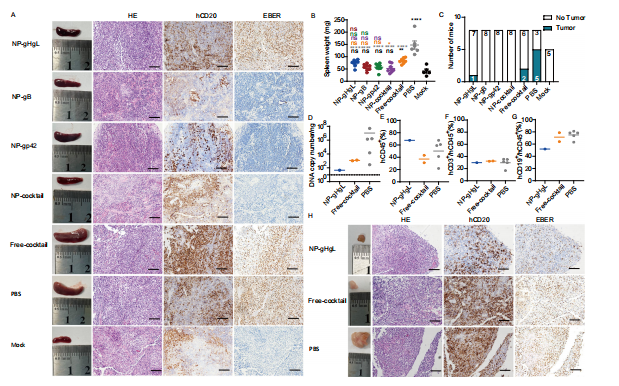
图7: 注射了由NP鸡尾酒诱导的IgG的人源化小鼠未出现EBV相关淋巴瘤
图(A)展示了代表性脾脏及其切片,这些切片经过苏木精和伊红(HE)染色、hCD20+免疫染色以及EBV编码RNA(EBER)原位杂交处理。每张图片代表一个实验组(比例尺= 50微米)(NP-gHgL,NP-gB,NP-gp42,NP-鸡尾酒和Free-鸡尾酒:n = 8;PBS,n = 6;Mock,n = 5)。B每个实验组(NP-gHgL,NP-gB,NP-gp42,NP-鸡尾酒和Free-鸡尾酒:n = 8;PBS,n = 6;Mock,n = 5)中个体小鼠在实验结束时或安乐死前(体重减少超过20%)的脾脏重量。由于样本腐烂,PBS组中两只小鼠在第7天和第10天死亡的样本未被收集。数据以平均值±标准误的形式展示。统计分析采用了一元方差分析(ANOVA)并结合了Turk多重比较检验,精确的P值见源数据文件。星号或ns的颜色表示统计学差异。
图(C)在NP-gHgL、自由鸡尾酒和PBS IgG处理组中,分别有1/8、2/8和5/8的小鼠发展出了EBV相关的淋巴瘤,这一诊断基于组织学观察。每个肿瘤的DNA拷贝数(D)以及hCD45+ (E)、hCD3+ (F)、hCD19+ (G)细胞的比例均被记录。水平线代表各组的平均值,虚线则表示检测限。(NP-gHgL组,n = 1;自由鸡尾酒组,n = 2;PBS组,n = 5)。H图展示了代表性的大体肿瘤和肿瘤组织(NP-gHgL组,n = 1;自由鸡尾酒组,n = 2;PBS组,n = 5),这些样本经HE染色、EBER杂交,并在尸检时对hCD20+进行了免疫染色(比例尺= 50微米)。源数据文件已提供。
创新性: 首次将EBV核心融合糖蛋白(gHgL/gB/gp42)与纳米载体结合,实现多靶点协同免疫。
转化意义: 鸡尾酒纳米疫苗为EBV预防性疫苗的临床开发提供了高效候选方案,尤其适用于高风险人群(如青少年和免疫缺陷患者)。
通用潜力: 该纳米平台可扩展至其他疱疹病毒疫苗设计。
常见细胞污染类型如何辨别及预防解决方法:细胞培养中常见的生物污染类型有7种,分别是细菌污染,支原体污染,原虫污染,黑胶虫污染,真菌污染,病毒污染以及非细胞污染,真菌污染来源,一般是来自实验服,并且具有气候性,多雨······
细胞聚团的原因分析及如何避免:培养物中细胞可能聚集的一些原因包括:1.过度消化、2.环境压力、3.组织分解、4.过度生长、5.污染等;如何避免聚团细胞的生成;首先确认当前细胞生长密度及状态,80%左右的生长密度即可进行······
细胞有空泡原因分析及解决方法:出现细胞空泡情况有1.细胞老化2.培养液错误配制;3.细胞消化时操作不当;4.污染等等,如细胞老化,解决方法,原代细胞使用较低代次进行实验,传代细胞避免传代次数过高···
细胞半换液和全换液操作步骤:第一种方式:细胞全换液;如果是贴壁细胞,可以用全量换液法,直接吸去全部旧培养基,补充足量新鲜完全培养基;第二种方式:细胞半换液;"细胞半换液"又称"细胞半量换液",即弃掉一半旧的培养基,再······
细胞生长缓慢的可能原因有哪些:细胞培养外部因素包括细胞培养基的配方和质量问题,培养条件不理想,污染问题,细胞自身因素包含细胞的健康状态,细胞密度过高或过低,细胞老化现象,细胞特性,当细胞生长出现缓慢的问题时,我······
常用胰腺癌细胞株动物模型及胰腺癌细胞株有哪些:胰腺癌研究中常用的动物模型主要包括化学物质诱导胰腺癌动物模型,基因工程胰腺癌小鼠模型和胰腺癌移植模型,常用的胰腺细胞株MIA-PACA-2人胰腺癌细胞,Capan-2人胰腺癌细······
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
上一篇:检测新突破3步搞定尼帕病毒抗体筛查!这种 "混合-读取" 技术让高危病毒监测更简单
下一篇:科研新发现查尔酮类化合物竟能 “熄灭” 萤火虫荧光?环结构决定生物发光抑制力
厦门爱恪信生物科技有限公司
手机:15859239971
邮箱:2205839769@qq.com
地址:厦门翔安火炬高新区翔星路96号建业楼D座602

微信公众号
ATCC细胞培养

技术支持
15859239971
Copyright©厦门爱恪信 闽ICP备19027235号-7
公安备案: XML地图
XML地图




 客服QQ
客服QQ