常见细胞污染类型如何辨别及预防解决方法
常见细胞污染类型如何辨别及预防解决方法:细胞培养中常见的生物污染类型有7种,分别是细菌污染,支原体污染,原虫污染,黑胶虫污染,真菌污染,病毒污染以及非细胞污染,真菌污染来源,一般是来自实验服,并且具有气候性,多雨······

发布时间:2025-05-12 08:59:41 细胞资源库平台 访问量:490
荧光素酶报告基因系统是一种基于荧光素酶催化底物氧化反应产生生物发光的检测技术,广泛应用于细胞生物学研究。其中,萤火虫荧光素酶(firefly luciferase, Fluc)因其高灵敏度、宽线性检测范围(约7~8个数量级)以及较短的半衰期(在哺乳动物细胞中约为3小时,在植物细胞中约为3.5小时)而成为最常用的报告基因。其发光信号强度在酶浓度为10⁻¹⁶ mol/L至10⁻⁸ mol/L的范围内与酶活性呈线性关系,并且在理想条件下可检测到低至10⁻²⁰ mol/L的荧光素酶活性。此外,荧光素酶报告基因系统具有非放射性、检测快速、灵敏度高(比氯霉素乙酰转移酶CAT高100倍)等优点,特别适用于高通量筛选和活细胞检测。通过将荧光素酶报告基因载体转染至宿主细胞后,可利用荧光素酶检测系统灵敏且便捷地监测基因表达水平,已成为细胞生物学研究中的重要工具。
英文标题:Development of luciferase-based highly sensitive reporters that detect ER-associated protein biogenesis abnormalities
中文标题:基于萤火虫荧光素酶的高灵敏度报告基因的开发,用于检测内质网相关蛋白质生物合成异常
发表期刊:《iScience》
影响因子:4.6
作者单位:
1.Advanced Materials, Tohoku University, Sendai, Miyagi 980-8577, Japan
2.Institute for Research Initiatives, Nara Institute of Science and Technology, Ikoma, Nara 630-0192, Japan
3.Cell Biology Center, Institute of Integrated Research, Institute of Science Tokyo, Yokohama, Kanagawa 226-8501, Japan
作者信息:
Hiroshi Kadokura, Nanshi Harada, Satoshi Yamaki
内质网(ER)是分泌途径蛋白质生物合成的关键场所,其定位和二硫键形成对蛋白质功能至关重要。现有检测ER蛋白质异常的方法(如基于GFP的荧光探针)存在灵敏度低、光毒性强、数据波动大等问题。本文受细菌MalF-LacZ报告系统的启发,通过工程化改造萤火虫荧光素酶(FLuc),开发了一种高灵敏度、高重复性的报告系统,用于检测ER中蛋白质定位缺陷和二硫键形成异常。
本研究通过工程化改造萤火虫荧光素酶(FLuc),引入多个半胱氨酸突变(如V61C、L115C等),使其在ER氧化环境中因二硫键错误折叠而失活;当ER发生定位缺陷或二硫键形成异常时,FLuc恢复活性。通过优化信号肽(如calreticulin的N端40个氨基酸)增强ER定位效率,并添加N-糖基化位点和FLAG标签用于检测。采用双荧光素酶报告系统(以海肾荧光素酶RLuc为内参)减少实验误差。通过siRNA敲低SRP54(信号识别颗粒关键组分)、小分子抑制剂CADA处理(靶向CD4信号序列)以及BPA(Ero1α抑制剂)和LMF1过表达/敲低实验,分别验证了报告系统对ER定位缺陷、药物诱导的蛋白质转运抑制以及ER氧化还原动态的检测能力。
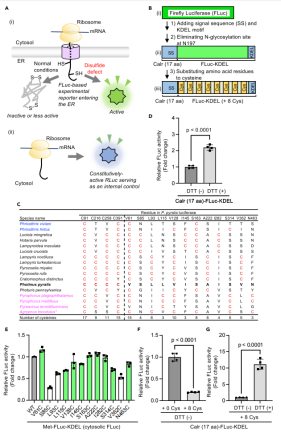
图1:工程化改造萤火虫荧光素酶以检测ER中的二硫键形成缺陷
研究团队通过在萤火虫荧光素酶(FLuc)中引入8个半胱氨酸突变(如V61C、L115C等),使其在ER的氧化环境中因异常二硫键形成而失活。
实验显示,正常条件下ER定位的FLuc活性显著降低(图1F),但用还原剂DTT处理细胞后,二硫键被破坏,FLuc活性恢复(图1G),表明突变成功赋予FLuc对ER氧化状态的敏感性。此外,通过对比野生型FLuc与突变体的活性(图1E),发现部分半胱氨酸替换不影响酶活性,为后续设计奠定了基础。
该结果表明,工程化的FLuc可作为检测ER二硫键形成缺陷的灵敏报告基因。
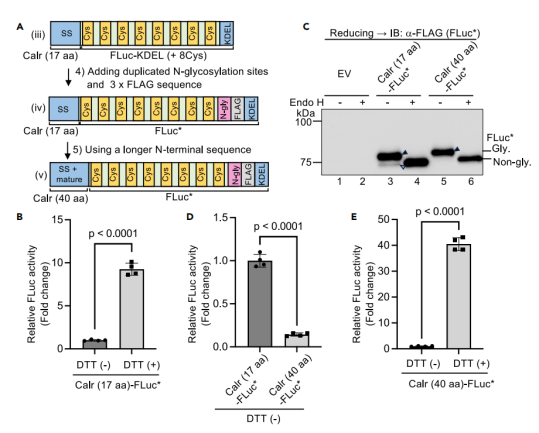
图2:优化信号肽设计增强ER定位并提高报告基因灵敏度
通过延长信号肽(从calreticulin的17个氨基酸扩展至40个氨基酸),并引入双N-糖基化位点,显著提高了FLuc*(突变体)的ER定位效率(图2C)。
原始设计(Calr(17 aa)-FLuc*)因部分蛋白滞留胞质导致背景活性较高,而优化后的Calr(40 aa)-FLuc几乎完全定位于ER(图2C)。
DTT处理后,Calr(40 aa)-FLuc活性提升更显著(图2E),表明信号肽优化减少了胞质“泄漏”,增强了对ER氧化缺陷的检测灵敏度。
这一改进为后续实验提供了高信噪比的报告系统。
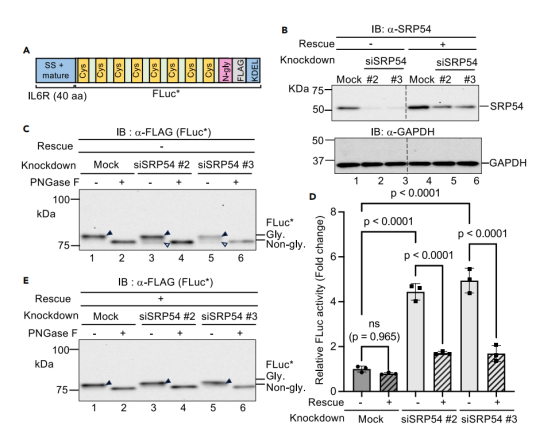
图3:SRP54敲低诱导的蛋白质ER定位缺陷检测
通过siRNA敲低信号识别颗粒(SRP)关键组分SRP54,验证报告系统对ER定位缺陷的检测能力。
SRP54敲低导致IL6R(40 aa)-FLuc部分未能进入ER(非糖基化形式累积,图3C),FLuc活性显著升高(图3D)。
而共表达siRNA抗性SRP54可回补缺陷(图3E),证明活性升高源于SRP依赖性ER定位受损,而非脱靶效应。该实验表明,FLuc报告系统能灵敏反映蛋白质靶向ER的异常,适用于研究分泌途径的调控机制。
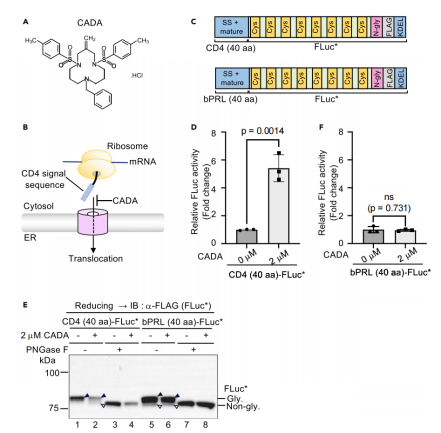
图4:基于CD4信号序列的报告系统检测病毒受体抑制剂CADA
将HIV受体CD4的信号序列(40个氨基酸)与FLuc融合,验证小分子抑制剂CADA对ER定位的特异性抑制。CADA处理导致CD4(40 aa)-FLuc活性显著升高(图4D),并伴随非糖基化蛋白累积(图4E),而对照信号序列(bPRL)无此现象(图4F)。此结果证明,FLuc*系统可定制化用于检测特定信号序列的功能异常,为高通量筛选靶向ER定位的抑制剂(如抗病毒药物)提供了新方法。
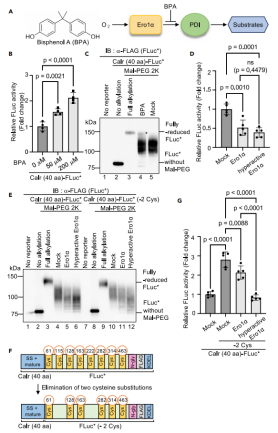
图5:Ero1α活性调控ER氧化还原环境的动态检测
通过抑制Ero1α(使用BPA)或过表达其超活性突变体(C104A/C131A),研究ER氧化还原状态变化。BPA处理剂量依赖性升高FLuc活性(图5B),而超活性Ero1α进一步降低活性(图5D),表明Ero1α是调控ER氧化能力的关键因子。通过Mal-PEG烷基化实验(图5C),直接观察到BPA处理导致FLuc二硫键减少(还原态比例增加)。此外,减少FLuc的半胱氨酸数目(Calr(40 aa)-FLuc(-2 Cys))可扩大检测范围,区分Ero1α野生型与突变体的氧化能力差异(图5G)。这些结果揭示了ER氧化系统的动态平衡及报告系统的灵活性。
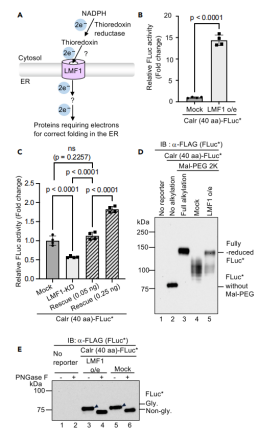
图6:LMF1通过调控还原当量供应影响ER氧化还原稳态
LMF1(脂酶成熟因子1)过表达显著升高FLuc活性(图6B),而敲低LMF1则降低活性(图6C),表明LMF1通过向ER提供还原当量维持其还原环境。Western blot显示,LMF1过表达导致FLuc二硫键减少(图6D),且不影响其ER定位(图6E)。此结果首次在非应激条件下证实LMF1对ER氧化还原稳态的直接调控,支持其通过跨膜电子传递参与蛋白质折叠的模型,并凸显FLuc*系统在揭示复杂调控网络中的高灵敏性。
本研究成功开发了基于FLuc的高灵敏度报告系统,可实时检测ER中蛋白质定位和二硫键形成异常,其灵敏度和重复性显著优于传统荧光探针。该系统不仅能识别病毒受体合成抑制剂(如CADA)诱导的定位缺陷,为药物筛选提供新工具,还揭示了LMF1通过调控ER氧化还原稳态(提供还原当量)影响蛋白质折叠的分子机制。通过调整FLuc*的半胱氨酸数量,可定制报告系统以检测ER氧化或还原状态的变化,为研究分泌蛋白生物合成机制及疾病相关靶点开发提供了重要技术平台。
常见细胞污染类型如何辨别及预防解决方法:细胞培养中常见的生物污染类型有7种,分别是细菌污染,支原体污染,原虫污染,黑胶虫污染,真菌污染,病毒污染以及非细胞污染,真菌污染来源,一般是来自实验服,并且具有气候性,多雨······
细胞聚团的原因分析及如何避免:培养物中细胞可能聚集的一些原因包括:1.过度消化、2.环境压力、3.组织分解、4.过度生长、5.污染等;如何避免聚团细胞的生成;首先确认当前细胞生长密度及状态,80%左右的生长密度即可进行······
细胞有空泡原因分析及解决方法:出现细胞空泡情况有1.细胞老化2.培养液错误配制;3.细胞消化时操作不当;4.污染等等,如细胞老化,解决方法,原代细胞使用较低代次进行实验,传代细胞避免传代次数过高···
细胞半换液和全换液操作步骤:第一种方式:细胞全换液;如果是贴壁细胞,可以用全量换液法,直接吸去全部旧培养基,补充足量新鲜完全培养基;第二种方式:细胞半换液;"细胞半换液"又称"细胞半量换液",即弃掉一半旧的培养基,再······
细胞生长缓慢的可能原因有哪些:细胞培养外部因素包括细胞培养基的配方和质量问题,培养条件不理想,污染问题,细胞自身因素包含细胞的健康状态,细胞密度过高或过低,细胞老化现象,细胞特性,当细胞生长出现缓慢的问题时,我······
常用胰腺癌细胞株动物模型及胰腺癌细胞株有哪些:胰腺癌研究中常用的动物模型主要包括化学物质诱导胰腺癌动物模型,基因工程胰腺癌小鼠模型和胰腺癌移植模型,常用的胰腺细胞株MIA-PACA-2人胰腺癌细胞,Capan-2人胰腺癌细······
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
上一篇:让耗竭T细胞‘发光’!新型小鼠模型助力破解癌症免疫治疗难题
下一篇:低氧肿瘤的免疫密钥被发现!这种蛋白让抗癌疫苗效果翻倍
厦门爱恪信生物科技有限公司
手机:15859239971
邮箱:2205839769@qq.com
地址:厦门翔安火炬高新区翔星路96号建业楼D座602

微信公众号
ATCC细胞培养

技术支持
15859239971
Copyright©厦门爱恪信 闽ICP备19027235号-7
公安备案: XML地图
XML地图




 客服QQ
客服QQ