常见细胞污染类型如何辨别及预防解决方法
常见细胞污染类型如何辨别及预防解决方法:细胞培养中常见的生物污染类型有7种,分别是细菌污染,支原体污染,原虫污染,黑胶虫污染,真菌污染,病毒污染以及非细胞污染,真菌污染来源,一般是来自实验服,并且具有气候性,多雨······

发布时间:2026-06-07 17:00:00 细胞资源库平台 访问量:37
英文标题:Peritumoural adipose tissue drives immune evasion in colorectal cancer via adipose-mesenchymal transformation
中文标题:肿瘤周围脂肪组织通过脂肪-间充质转化驱动结直肠癌免疫逃逸
发表期刊:《Nature Cell Biology》
影响因子:19.1
作者单位:
中山大学肿瘤防治中心
中山大学等
作者信息:
Jin-Hong Wang, Yong-Qiang Zheng, Zheng-Yu Qian, Huai-Qiang Ju等
研究背景:
1.临床/生物学问题:
结直肠癌(CRC)周围常存在大量内脏脂肪组织(tVAT),但其在肿瘤免疫逃逸中的具体作用尚不明确。
2.现有研究的局限性:
多数研究聚焦于肿瘤内部的微环境(TME),忽视了肿瘤周围脂肪组织对免疫系统的调控作用。
3.研究切入点与创新点:
本研究首次通过单细胞RNA测序(scRNA-seq)系统描绘了tVAT的免疫图谱,揭示其通过CXCL12-CXCR4轴竞争性“截留”免疫细胞,促进肿瘤免疫逃逸,并提出“脂肪-间充质转化”(Adipose-Mesenchymal Transformation)这一新概念。
1.动物模型构建:
使用C57BL/6J小鼠、BALB/c小鼠、NSG小鼠等,建立MC38、CT26、E0771等皮下及盲肠原位肿瘤模型。
2.细胞培养与处理:
使用MC38、CT26、E0771细胞系,以及小鼠脂肪来源干细胞(ASCs),进行肿瘤条件培养基(CM)刺激、TGF-β1诱导分化等实验。
3.单细胞与单核RNA测序:
对tVAT、dVAT、肿瘤、正常组织进行scRNA-seq和snRNA-seq,分析细胞亚群、轨迹发育、TCR/BCR克隆性。
4.免疫组化与多重免疫荧光:
使用mIHC、IF、H&E染色验证TLS结构、adCAF标志物(FAP、PDGFRB、MDK)及免疫细胞浸润。
5.功能性实验:
细胞趋化实验(3D Chemotaxis Assay)
T细胞共培养与激活实验
流式细胞术检测免疫细胞亚群及CXCR4表达
ELISA检测CXCL12、TGF-β1分泌
基因敲除与细胞清除模型:
Cxcl12条件性敲除小鼠(Cxcl12fl/fl)
Mdk-DTR小鼠特异性清除adCAF
ATS-GNP纳米颗粒特异性消除脂肪组织
6.临床样本分析:
纳入67例局部晚期CRC患者,评估tVAT面积与免疫治疗响应之间的相关性。
图1 | 单细胞RNA图谱揭示了tVAT的组织特异性免疫景观。
a. 实验设计示意图,展示了从结直肠癌(CRC)患者身上采集肿瘤、肿瘤周围内脏脂肪组织(tVAT)和远端内脏脂肪组织(dVAT)进行单细胞RNA测序(scRNA-seq)分析的流程(使用BioRender.com绘制)。
b. 综合UMAP图,展示了70个肿瘤样本、12个正常样本、12个tVAT样本和12个dVAT样本中总共371,381个高质量单细胞的转录图谱,并根据经典标记基因鉴定出13种不同的细胞亚群。
c. 热图显示了四种组织类型(tVAT、dVAT、肿瘤、正常)中主要细胞亚群的相对比例。
d. 使用Milo方法进行差异丰度分析,识别出tVAT(n=12)和dVAT(n=12)之间细胞邻域(Nhoods)的差异。节点大小与每个Nhood中的细胞数量成正比,图边表示相邻Nhood之间共享的细胞数量。显示为白色的节点表示差异不显著(假发现率 ≥ 0.1)。
e. Beeswarm图展示了tVAT(n=12)和dVAT(n=12)之间所有细胞类型的Nhoods丰度调整后的对数2倍变化(log2 FC)分布。
f. 箱线图比较了tVAT(n=12)和dVAT(n=12)中CD4+ T细胞、CD8+ T细胞、B细胞和基质细胞的相对丰度(使用双侧配对Wilcoxon检验)。箱线从下到上表示第一四分位数、中位数、第三四分位数,须线表示全距。
g. CRC患者dVAT和tVAT切片中三级淋巴结构(TLSs)的代表性苏木精-伊红(H&E)染色(23个样本)和多重免疫组化(mIHC)染色(4个样本)图像。比例尺,200 μm。
图2 | tVAT与dVAT中淋巴细胞的特性比较。
a. UMAP图展示了CD4+ T细胞、CD8+ T细胞/自然杀伤(NK)细胞和B/浆细胞的转录异质性及其亚群。
b. 热图显示了不同淋巴细胞亚群在tVAT、dVAT、肿瘤和正常组织中的组织分布偏好。
c. Beeswarm图展示了tVAT(n=12)和dVAT(n=12)之间淋巴细胞亚群的差异丰度。
d. 单个患者的dVAT、肿瘤和tVAT中前20个扩增最多的T细胞受体(TCR)克隆型的纵向追踪。每个色带代表一个独特的TCR克隆型,列之间的彩色条带表示不同组织间共享的克隆型。
e. 箱线图比较了tVAT(n=3)和dVAT(n=3)中TCR克隆多样性的差异(使用双侧配对Student's t检验)。
f. 热图显示了各组织样本间及其与对应肿瘤之间B细胞受体(BCR)克隆型的重叠情况。
g. 箱线图比较了tVAT(n=3)和dVAT(n=3)与各自肿瘤之间的BCR克隆型重叠度(使用双侧配对Student's t检验)。
图3 | tVAT与肿瘤组织竞争浸润性淋巴细胞。
a. MC38-OVA荷瘤C57BL/6J小鼠(每组n=5)在假手术、对照组或PAT切除后的代表性肿瘤图像(左)和肿瘤生长曲线(右)。
b. CT26荷瘤BALB/c小鼠(每组n=5)在假手术、对照组或PAT切除后的代表性肿瘤图像(左)和肿瘤生长曲线(右)。
c. 特异性减少PAT的ATS-GNP纳米颗粒(含CaCO3核和PLG壳,偶联ATS肽)的作用机制示意图。
d. C57BL/6J、BALB/c-Nude和NSG小鼠中MC38肿瘤在假手术或PAT切除后的生长曲线(每组n=5)。
e. C57BL/6J小鼠中E0771肿瘤在假手术、对照(切除对侧脂肪)或PAT切除后第16天的代表性图像(每组n=5)。
f. C57BL/6J小鼠中E0771肿瘤在假手术、对照或PAT切除后第16天的肿瘤重量(每组n=5)。
g. MC38-OVA肿瘤中活细胞门控的CD45+细胞代表性流式细胞术图。
h. 小提琴图展示了CRC患者所有细胞类型中CXCR7的表达水平(使用Kruskal-Wallis检验)。
i. 构建Cxcl12条件性敲除(cKO)小鼠的编辑策略。
j. 在PAT附近构建对照和Cxcl12fl/fl cKO小鼠MC38-OVA肿瘤的实验设计。
k. 蛋白质印迹(Western blot)验证对照和Cxcl12fl/fl cKO小鼠PAT中Cxcl12的敲除效率。
l. RT-qPCR验证对照和Cxcl12fl/fl cKO小鼠(n=6)的肝脏、脾脏、子宫和肿瘤组织中的Cxcl12敲除效率。
图4 | CXCL12-CXCR4信号轴介导tVAT中淋巴细胞的保留。
a. 基于配体-受体对(前六对)分析tVAT中基质细胞与淋巴细胞之间的细胞间通讯(左),并比较tVAT、dVAT和肿瘤中的通讯概率(右)。
b. 标记图显示了tVAT、dVAT和肿瘤中CD8+ T细胞、CD4+ T细胞、B细胞、浆细胞与基质细胞群之间显著的CXCL12-CXCR4相互作用。线条宽度代表通讯概率。
c. 小提琴图展示了CRC患者所有细胞类型中CXCL12(上)和CXCR4(下)的表达水平。
d. 小提琴图比较了CRC患者dVAT与tVAT(上)以及肿瘤与tVAT(下)中CXCL12的表达水平(使用双侧Wilcoxon检验)。
e. PAT C57BL/6J小鼠模型中,使用IgG或抗CXCL12抗体治疗(左)的实验设计,以及第16天实验结束时的代表性MC38肿瘤图像(右)(每组n=5)。
f. C57BL/6J小鼠(n=5)中MC38肿瘤在实验第16天的肿瘤生长曲线(左)和肿瘤重量(右)。
g. 对照和Cxcl12fl/fl cKO小鼠(n=6)中MC38肿瘤的代表性图像(左)和肿瘤生长曲线(右)。
h. 对照和Cxcl12fl/fl cKO小鼠(n=6)在实验第16天的MC38肿瘤重量。
i. 流式细胞术分析对照和Cxcl12fl/fl cKO小鼠(n=6)MC38肿瘤中不同CXCR4+免疫细胞的浸润数量。
j. 使用T细胞作为“传感器”、条件培养基作为“汇”的趋化实验示意图(左),以及对照或CXCL12诱导的T细胞迁移1小时的轨迹图(右)。
k. 通过流式细胞术定量分析,在有或没有PAT切除(或切除对侧腹股沟脂肪作为对照)的情况下,MC38肿瘤中CXCR4+ CD45.1+ T细胞的绝对数量(每组n=5)。
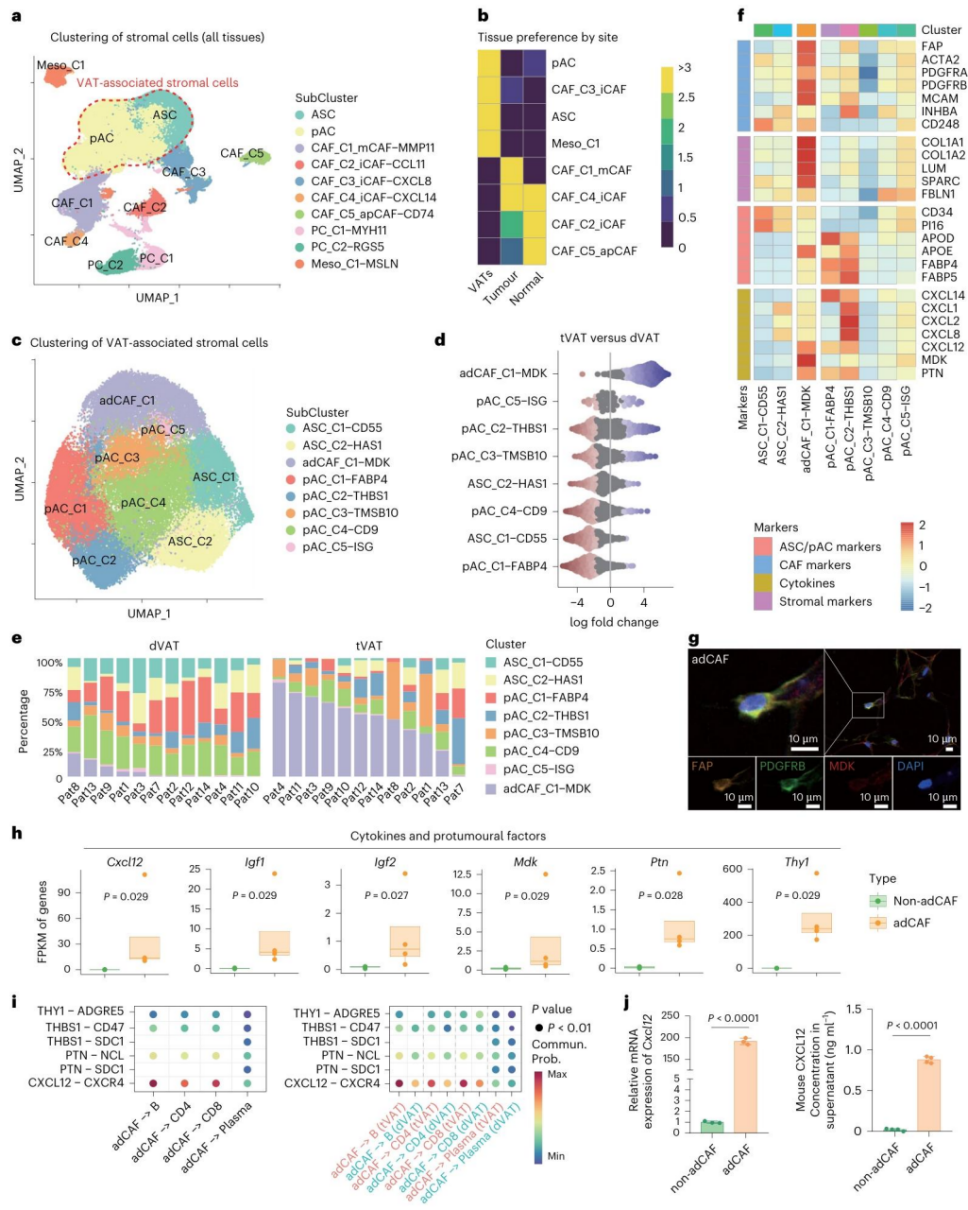
图5 | 肿瘤在tVAT中诱导脂肪-间充质转化。
a. CRC患者tVAT、dVAT、肿瘤和正常组织中所有基质细胞的UMAP图,根据推断的细胞类型标记了10个聚类(ASCs、pACs、CAFs、周细胞PCs、间皮细胞Mesos)。
b. 热图展示了八种基质细胞亚型在不同组织类型中的分布。
c. CRC患者tVAT和dVAT中八种VAT相关基质细胞亚型的UMAP图,包括ASC、pAC和adCAF。
d. Beeswarm图展示了tVAT和dVAT之间VAT相关基质细胞类型在Nhoods中的分布和丰度。
e. 堆叠柱状图展示了八种VAT相关基质细胞亚型在dVAT和tVAT中的丰度。
f. 热图显示了VAT相关基质细胞类型中多种标记基因的RNA表达,包括ASC/pAC标记、CAF标记、细胞因子和基质标记。
g. CRC患者tVAT样本中adCAF存在的代表性多重免疫荧光图像。比例尺,10 μm。DAPI(4',6-二脒基-2-苯基吲哚)用于染核。
h. 小鼠异种移植模型PAT中,富集adCAF的基质细胞(n=4)和非adCAF基质细胞(n=4)中多种细胞因子和促肿瘤因子的相对表达水平。数据以箱线图呈现(箱线边界为第一至第三四分位数,底部和顶部须线为最小至最大值,中心线为中位数)。
i. 基于配体-受体对(前六对)分析tVAT中adCAF与淋巴细胞之间的细胞间通讯(左),以及tVAT与dVAT之间的比较(右)。
j. RT-qPCR(左)和ELISA(右)检测小鼠PAT中分选的富集adCAF的基质细胞和非adCAF基质细胞中CXCL12的RNA表达和蛋白分泌水平。
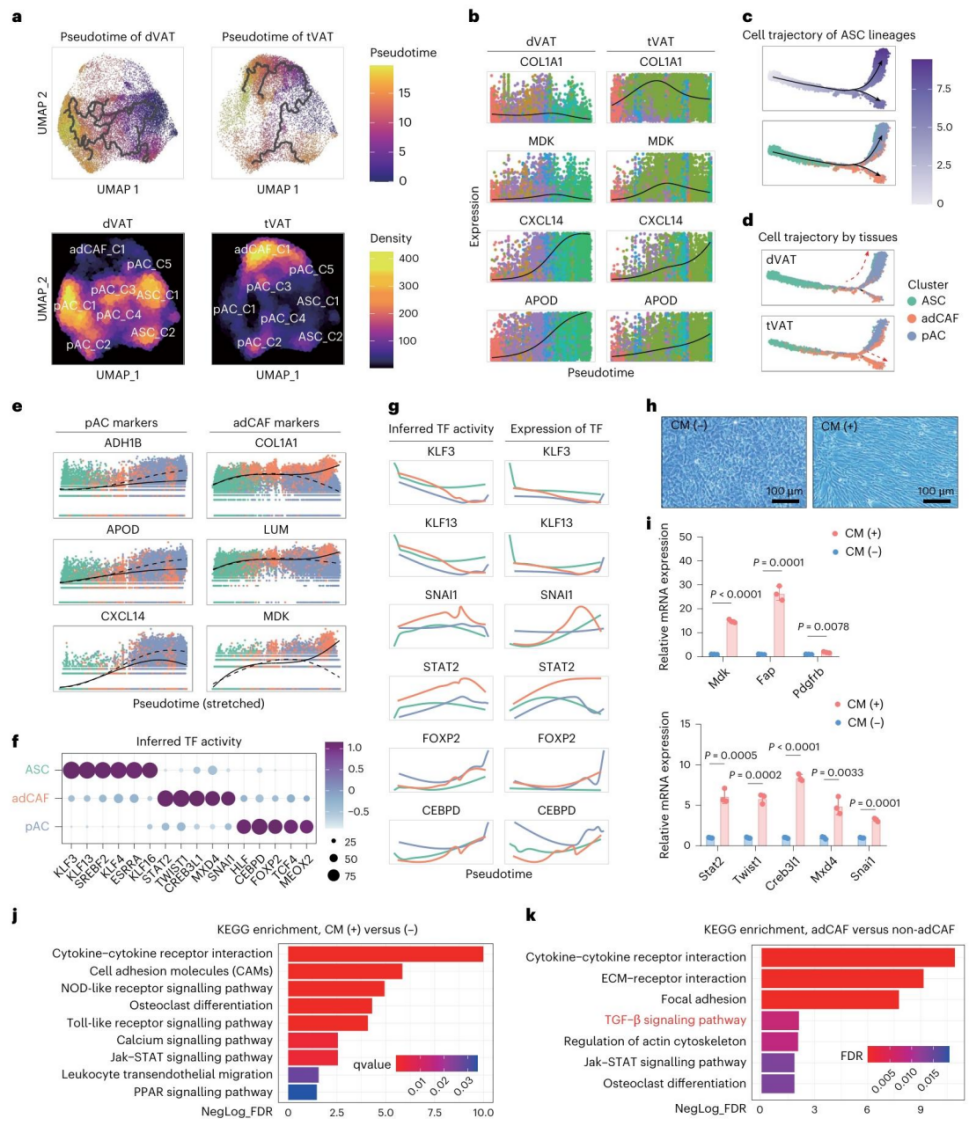
图6 | 肿瘤来源的因子诱导ASC分化为adCAF。
a. Monocle3推断的ASC来源的pAC和ASC来源的adCAF的细胞轨迹(上)和细胞丰度(下),展示了dVAT(左)和tVAT(右)中的细胞轨迹。伪时间颜色代码如右框所示。
b. Monocle3伪时间分析显示,多个标记基因在dVAT(左)和tVAT(右)中随时间的动态表达变化。
c. Monocle2推断的ASC来源的pAC和ASC来源的adCAF的伪时间分析(上)和细胞轨迹(下)。伪时间颜色代码如右框所示。
d. Monocle2推断的ASC来源的pAC和ASC来源的adCAF在dVAT(上)和tVAT(下)中的细胞轨迹。
e. Monocle2伪时间分析显示pAC标记物(左)和adCAF标记物(右)随时间的动态表达变化。
f. 点图显示了ASC、pAC和adCAF中推断的差异转录因子(TF)活性。
g. 热图显示了ASC、pAC和adCAF中转录因子推断活性和RNA表达水平的动态变化。
h. 使用MC38肿瘤来源的条件培养基(CM)处理72小时后,小鼠ASC(mASC)形态的代表性图像。比例尺,100 μm。
i. RT-qPCR检测CM处理的mASC中adCAF相关基因(上)和转录因子(下)的RNA表达水平。
j. KEGG通路富集分析条形图,显示肿瘤CM处理的mASC中显著富集的信号通路。
k. KEGG通路富集分析条形图,显示富集adCAF的基质细胞与非adCAF基质细胞之间差异富集的信号通路。
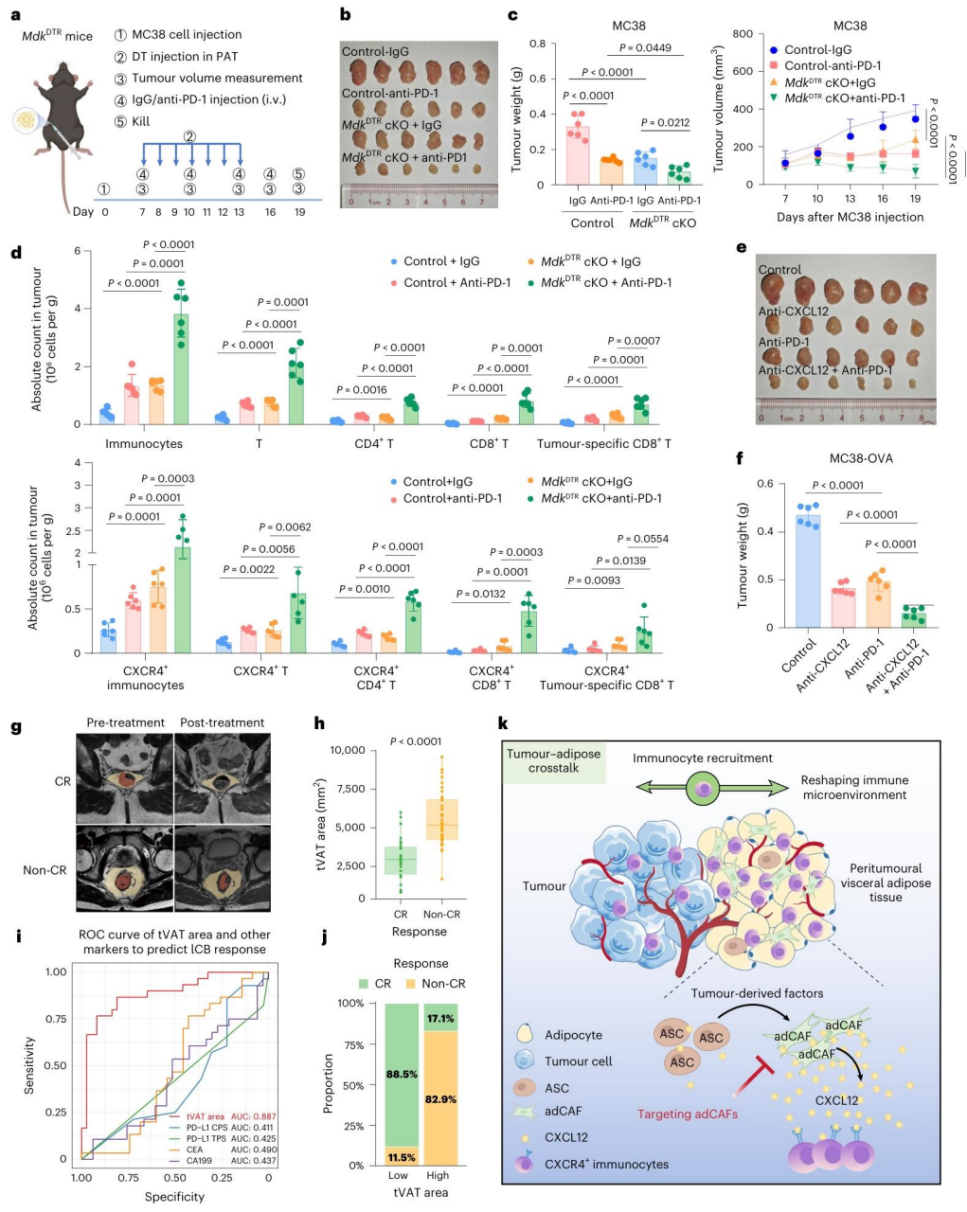
图7 | 肿瘤-脂肪组织相互作用的临床意义。
a. 在PAT中使用Mdk-DTR系统特异性清除adCAF并结合抗PD-1治疗的实验设计(每组n=6)b. 实验结束时(第16天),来自对照组(同窝对照)、抗PD-1单药组、Mdk-DTR(DT处理)组和联合治疗组的代表性MC38-OVA肿瘤图像。
c. (左图)各组小鼠的个体肿瘤生长曲线和(右图)肿瘤重量比较(每组n=6)。
d. 流式细胞术分析各组MC38-OVA肿瘤中免疫细胞(T细胞、CD4+ T、CD8+ T、肿瘤特异性CD8+ T)和CXCR4+免疫细胞亚群的浸润绝对数量(每组n=6)。
e. 接受新辅助免疫-放化疗的pMMR局部晚期CRC患者的治疗反应评估流程图。
f. 箱线图比较达到完全缓解(CR)和未达到完全缓解(非CR)患者的治疗前PAT面积(n=67)。数据以箱线图呈现(箱线边界为第一至第三四分位数,底部和顶部须线为最小至最大值,中心线为中位数)。
g. 箱线图分别比较T3期和T4期患者中CR与非CR组的tVAT面积差异。
h. ROC曲线分析,比较治疗前PAT面积与传统指标(CPS、TPS、CEA、CA199)对pMMR CRC患者免疫-放化疗反应的预测能力(n=67),并显示最佳截断值。
i. 根据最佳截断值,比较tVAT高组和低组之间的病理完全缓解(pCR)率。
j. 图形摘要,描绘了肿瘤如何重塑tVAT中的基质环境,以及tVAT如何通过与肿瘤竞争免疫细胞来促进免疫逃逸的模型(使用BioRender.com绘制)。
本研究首次系统揭示了结直肠癌周围脂肪组织(tVAT)通过“脂肪-间充质转化”形成adCAF,分泌CXCL12,竞争性截留CD8+ T细胞,促进免疫逃逸。靶向CXCL12-CXCR4轴或清除adCAF可显著增强抗PD-1治疗疗效。此外,tVAT面积可作为预测免疫治疗响应的临床生物标志物。
文献意义:
1.首次提出“脂肪-间充质转化”概念,揭示tVAT在肿瘤免疫调控中的新机制。
2.明确adCAF为CXCL12的主要来源,拓展了CAF的功能认知。
3.提供tVAT作为免疫治疗靶点及影像学生物标志物的临床证据。
4.为结直肠癌及其他腹部肿瘤的免疫治疗增敏提供了新策略。
常见细胞污染类型如何辨别及预防解决方法:细胞培养中常见的生物污染类型有7种,分别是细菌污染,支原体污染,原虫污染,黑胶虫污染,真菌污染,病毒污染以及非细胞污染,真菌污染来源,一般是来自实验服,并且具有气候性,多雨······
细胞聚团的原因分析及如何避免:培养物中细胞可能聚集的一些原因包括:1.过度消化、2.环境压力、3.组织分解、4.过度生长、5.污染等;如何避免聚团细胞的生成;首先确认当前细胞生长密度及状态,80%左右的生长密度即可进行······
细胞有空泡原因分析及解决方法:出现细胞空泡情况有1.细胞老化2.培养液错误配制;3.细胞消化时操作不当;4.污染等等,如细胞老化,解决方法,原代细胞使用较低代次进行实验,传代细胞避免传代次数过高···
细胞半换液和全换液操作步骤:第一种方式:细胞全换液;如果是贴壁细胞,可以用全量换液法,直接吸去全部旧培养基,补充足量新鲜完全培养基;第二种方式:细胞半换液;"细胞半换液"又称"细胞半量换液",即弃掉一半旧的培养基,再······
细胞生长缓慢的可能原因有哪些:细胞培养外部因素包括细胞培养基的配方和质量问题,培养条件不理想,污染问题,细胞自身因素包含细胞的健康状态,细胞密度过高或过低,细胞老化现象,细胞特性,当细胞生长出现缓慢的问题时,我······
常用胰腺癌细胞株动物模型及胰腺癌细胞株有哪些:胰腺癌研究中常用的动物模型主要包括化学物质诱导胰腺癌动物模型,基因工程胰腺癌小鼠模型和胰腺癌移植模型,常用的胰腺细胞株MIA-PACA-2人胰腺癌细胞,Capan-2人胰腺癌细······
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
上一篇:基于新型G系列腔肠素变体的NanoLuc驱动多重成像平台
下一篇:MicroRNA-141通过EGFR/β-catenin信号通路诱导自噬调控肺缺血再灌注损伤
厦门爱恪信生物科技有限公司
手机:15859239971
邮箱:2205839769@qq.com
地址:厦门翔安火炬高新区翔星路96号建业楼D座602

微信公众号
ATCC细胞培养

技术支持
15859239971
Copyright©厦门爱恪信 闽ICP备19027235号-7
公安备案: XML地图
XML地图




 客服QQ
客服QQ