常见细胞污染类型如何辨别及预防解决方法
常见细胞污染类型如何辨别及预防解决方法:细胞培养中常见的生物污染类型有7种,分别是细菌污染,支原体污染,原虫污染,黑胶虫污染,真菌污染,病毒污染以及非细胞污染,真菌污染来源,一般是来自实验服,并且具有气候性,多雨······

发布时间:2023-10-24 11:43:35 细胞资源库平台 访问量:411
今天推荐的是由科罗拉多大学健康科学中心牙科学院基础科学和口腔研究系在1997年1月18日发表于In Vitro Cellular & Developmental Biology-Animal(2021IF:2.1001,JCRQ1)的一篇文章,通讯作者是J.T.TURNER教授,研究主要阐述了SV40永久化大鼠腮腺腺癌细胞株的开发与特征描述。
使用含有猿猴病毒(SV40)基因组的质粒,通过CaPO4沉淀法转染大鼠腮腺唾液腺凋亡细胞。在30个克隆细胞系中,有2个细胞系被证明具有中度细胞分化和唾液腺胰腺细胞功能。对这两种细胞系进行的功能研究表明,肾上腺素能激动剂(异丙肾上腺素)、血管活性肠肽前列腺素E和福斯可林能有效促进细胞内环腺苷3':5'-环单磷酸的产生。苯肾上腺素、氨甲酰胆碱能有效增加磷酸肌醇的产生和细胞内游离钙的水平,而物质则会影响细胞内游离钙的产生。通过间接免疫荧光分析,两种细胞系都表达了SV40大T细胞。电子显微镜评估记录了中高水平的细胞分化,包括维持连接复合体、细胞极化、存在适量分泌颗粒。两种细胞系的倍增时间分别为22小时和36小时。
过去几年中,在确定大鼠唾液腺尖突细胞无血清原代培养对营养、激素、氧气和生长的要求方面取得了重大进展。维持尖腺细胞细胞分化和正常分泌功能所必需的重要细胞外基质蛋白已经确定。在无血清条件下,在35%02和5%CO2的环境中,在含有层粘连蛋白的重组大鼠尾胶原凝胶上培养大鼠唾液腺尖头细胞,在培养的前15天,尖头细胞能够维持中等水平的细胞分化和功能。然而,到了第22天,细胞分化水平下降,出现了严重的细胞死亡。唾液腺研究的主要障碍之一是无法繁殖大量相同的、分化良好的尖腺细胞用于体外研究。建立具有高度细胞分化和接近正常尖状体功能的永生化细胞系对于许多唾液腺尖状体细胞和分子生物学的体外研究至关重要。在过去几年中,有几种不同的方法被用于开发永生化大鼠唾液腺细胞系(Patton和Wellner,1993年;Prasad等人,1994年)。年轻的成年大鼠腮腺唾液腺细胞在原代培养中生长。用pSVaneo质粒转染细胞,随后经过新霉素抗生素选择,培育出永生化细胞系。在本报告中,研究人员使用了以前用于建立人类上皮细胞系的pSVori-质粒构建体(Gruenert等人,1988年;Quissell,1997年)。研究人员并没有利用抗生素选择来鉴定单个细胞系,而是建立了一个选择性筛选来鉴定分化良好的克隆细胞系,以防止转染高分化尖顶细胞后可能出现的损失,因为高分化尖顶细胞对选择性抗生素的毒性更敏感。如果没有细胞外基质作为基质,未转染的成年大鼠胰腺细胞无法在体外存活,也无法在塑料上存活(Quissell等人,1994年)。因此,转染细胞在转染后的最初几个阶段都是在细胞外基质上维持的,以便细胞在塑料上生长之前有时间适应(Quissell等人,1997年)。此外,每个细胞系都是在转染后几个阶段才进行克隆的,这样可以让永生化细胞在进行克隆选择和在塑料上培养之前进一步适应和选择。
1.使用起源缺陷的SV40质粒转染大鼠腮腺泌乳素细胞
通过一系列实验转染大鼠腮腺腺泡细胞,分离出30个不同的细胞克隆;其中11个克隆细胞系细胞角蛋白1-8呈阳性,19个克隆细胞系波形蛋白呈阳性。后一种细胞系被舍弃。随后分析了剩余的11个细胞系对各种分泌物的[Ca2+]i迁移反应。
图1A是大鼠腮腺细胞在转染后两代和克隆前在丝裂霉素C处理过的3T3成纤维细胞上生长的相位对比显微照片。图1A给出了上皮细胞典型的鹅卵石模式,大多数转染细胞系都表现出这种模式。只有那些波形蛋白阳性(即来源于成纤维细胞)的细胞系没有表现出这种生长模式。这种鹅卵石外观甚至出现在上皮细胞系中,后来证明这些细胞系的分化程度很低。图1B是对转染后的大鼠腮腺细胞进行免疫荧光分析以检测SV40大T抗原的例子。转染和克隆后检测的所有细胞系在SV40大T抗原表达方面均呈阳性。如图1B所示,SV40大T抗原的水平因细胞而异,但由于SV40大T抗原的转化活性,SV40表达的主要亚细胞区是细胞核。
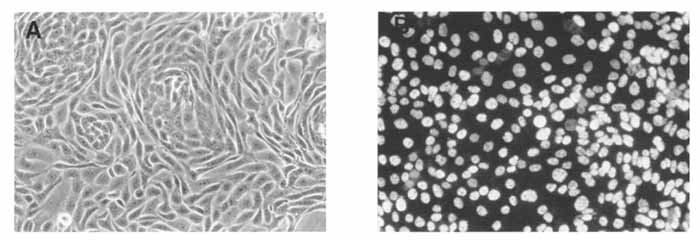
图1.使用起源缺陷的SV40质粒转染大鼠腮腺泌乳素细胞
2.初步筛选
在分离出克隆上皮细胞系后,研究人员利用筛选程序来确定并开始描述那些由分化良好的尖突细胞组成的细胞系。研究人员发现,通过分析[Ca2+]i的变化来确定单个克隆细胞对各种激动剂刺激的反应,是一种有效而快速的方法,可用于识别具有合理细胞和功能分化水平的细胞系。图2A展示了使用新鲜分散的大鼠腮腺唾液腺细胞进行典型研究的结果。这些细胞受到P物质刺激后,细胞内游离钙的变化幅度总是最大的。氨甲酰胆碱(一种毒蕈碱胆碱受体激动剂)和去甲肾上腺素(通过肾上腺素受体)的反应较低,但始终可以测量到,约为基础值的三至四倍。ATP在刚分散的细胞中产生的反应较小,但可重复。
在对单个细胞系进行永生化和鉴定之后,单个细胞系对这些激动剂的反应各不相同,而且差异极大。在测试的任何单个细胞系中,研究人员都无法证明由P物质受体引起的[Ca2+]i反应。许多上皮细胞系在胆碱能和/或ct肾上腺素能受体刺激后也没有表现出反应。只有两个克隆腮腺尖状腺细胞系(C5和C10)表现出明显的胆碱能和肾上腺素能受体介导的反应,如图2(B和C组)所示。除了对P物质的反应消失外,这两种细胞系对ATP的最大[Ca2+]i反应也急剧增加。
作为初步筛选程序的一部分,研究人员测定了细胞系对β-肾上腺素能受体异丙肾上腺素的反应性。几乎所有上皮细胞系对异丙肾上腺素的反应都是细胞内环磷酸腺苷水平的增被他加。不过,不同细胞系的反应程度差异很大。无论细胞系的分化程度如何,β-肾上腺素能受体偶联腺苷酸环化酶反应系统都保持完整。因此,β-肾上腺素能受体的活性似乎不是分化的鉴别指标。
为了评估每种细胞系的细胞分化程度,研究人员都进行了光镜和电子显微镜研究。那些没有对胆碱能和邻肾上腺素能激动剂表现出充分的受体介导的[Ca2+]i动员反应的上皮细胞系也表现出较低的细胞分化水平。在已建立的30个细胞系中,有2个细胞系根据形态和功能标准被证明具有合理的细胞分化水平。研究人员对这两种细胞系进行了以下研究:C5和C10。
![图2.使用H&L仪器观察分泌物对装载fura-2/AM的细胞中[Ca2+]i的影响](http://wh-nb4omimgoi1zr2ni4bb.my3w.com/static/upload/image/20231024/1698129293186055.jpg)
图2.使用H&L仪器观察分泌物对装载fura-2/AM的细胞中[Ca2+]i的影响
3.克隆腮腺腺泡细胞系(C5和C10)的表征
两种细胞系的细胞都生长在盖玻片上,然后浸泡在2.5%戊二醛和0.1M NaH2PO4 pH7.4中的0.1M蔗糖中90分钟,用缓冲液洗涤三次,在Millonigs缓冲液中的1%OsO4中后固定1小时,用乙醇和环氧丙烷脱水,包埋在Medcast中。通过光学显微镜观察(未图示),转染20次后固定的C5主要由两到四层丰满的细胞组成,其中含有散在的、密度适中的分泌颗粒。偶尔有细胞群形成微囊,伸入培养基中。C10在转染16个传代后固定,已形成由立方体和丰满细胞组成的单层细胞,偶尔含有中等密度的分泌颗粒。
细胞系C5和ClO的超微结构(图3-5)在许多方面都很相似。几乎所有细胞都含有数量适中的分泌颗粒,具有双相(电子致密和透明)亚结构和中等至大量粗糙的内质网,以及许多游离核糖体。外渗过程中的分泌颗粒很少见。这两个样本的细胞在内侧表面通过由紧密连接、中间连接(其中一些类似于间隙连接)和半球体组成的三方连接复合体相互连接。紧靠内侧质膜下方的末端网(由微丝组成的网状结构)连接着紧密连接。在有丝分裂过程中看到的所有细胞都含有分泌颗粒和大量粗糙的内质网。
C5和Cl0之间也有一些明显的差异。在C5中,几个细胞层之间有许多从内侧表面侵入的管腔状细胞间隙,由三部分连接复合体封闭,并镶有微绒毛。再看光镜下的微囊,面向空腔的细胞表面有钝突,由脱膜小体连接,而不是紧密连接。面向介质的细胞外表面则由紧密连接体连接,并镶有细长的微绒毛。这些观察结果表明,微囊腔是细胞外空间,已扩大到许多内衬细胞变薄的程度,这显然是液体积聚的结果。另一方面,ClO大多为单层,内侧表面没有管腔状的细胞间延伸,底部的细胞外基质要厚得多。
由于SpexCMlTllI双激发分光荧光仪的灵敏度和反应能力更强,因此研究人员使用它来进一步鉴定这两种克隆细胞系。图6显示了两种细胞系对不同浓度的毒蕈碱、胆碱能和P2-嘌呤能受体激动剂的反应。这些数据证实了研究人员最初的观察结果,即细胞在原代培养、转染和永生化后,P,介导的反应增强。如图6所示,两种细胞系对所测试的各种激动剂都给出了相似的剂量-反应模式。此外,两种细胞系对ATP和尿苷-5'-三磷酸(UTP)的剂量-反应曲线相等,这表明介导这些效应的嘌呤受体属于Pzu亚型,研究人员中的一位最近克隆并表达了这种受体,而且人类下颌下腺细胞系HSG-PA中也存在这种受体。
先前的研究表明,Pzu-嘌呤能、毒蕈碱能、胆碱能和a-肾上腺素能受体是大量细胞表面受体的代表,它们与肌醇磷脂特异性磷脂酶C功能耦合,导致许多细胞类型(包括唾液腺腺体细胞)内的钙动员。图7显示了这些受体在两种细胞系中增强肌醇磷酸盐生成的能力。这些数据与图6显示的结果一致,表明存在一个涉及IP的典型钙离子级联。
大鼠腮腺腺泡细胞中直接影响初级分泌液的产生和外泌的两个主要信号转导途径是钙信号途径和环AMP信号途径。尽管永生化细胞克隆对肾上腺素能再受体刺激的敏感性并不能很好地反映细胞的整体差异和功能,但环状AMP途径仍然是完全分化的唾液腺ac-inar细胞的重要信号转导途径。为了更全面地描述这两种细胞系中的环磷酸腺苷通路,并评估它们与各种细胞表面受体的联系,研究人员进行了以下研究。图8展示了一系列研究的结果,以评估肾上腺素能、血管活性肠肽和前列腺素E受体与腺苷酸环化酶的活化以及随后细胞内环状AMP的增加之间的耦合关系。-肾上腺素能受体刺激的反应最大,其次是前列腺素E(PGE)受体的激活,而对血管活性肠肽(VIP)受体的研究结果不一。虽然相对较小,但两种细胞系对VIP的反应是一致的。
在单个细胞系的筛选和生长过程中,研究人员发现许多细胞克隆在培养过程中的生长速度明显不同。图9显示了转染后两种细胞系的生长速度。细胞系C10在转染和再培养后经过一段时间的延迟,其加倍时间约为22小时,而细胞系C5的加倍时间约为36小时。

图3.C5的电子显微照片

图4.C5的电子显微照片

图5.C10的电子显微照片
![图6.在C5和Cl细胞中装入fura-2/AM,并测定[Ca2+]对指定浓度的UTP(开圆圈)、ATP(闭圆圈)和卡马胆碱(开三角形)的反应变化](http://wh-nb4omimgoi1zr2ni4bb.my3w.com/static/upload/image/20231024/1698129293509501.jpg)
图6.在C5和Cl细胞中装入fura-2/AM,并测定[Ca2+]对指定浓度的UTP(开圆圈)、ATP(闭圆圈)和卡马胆碱(开三角形)的反应变化

图7.促泌剂对腮腺尖锐湿疣细胞株中三磷酸肌醇总含量的影响
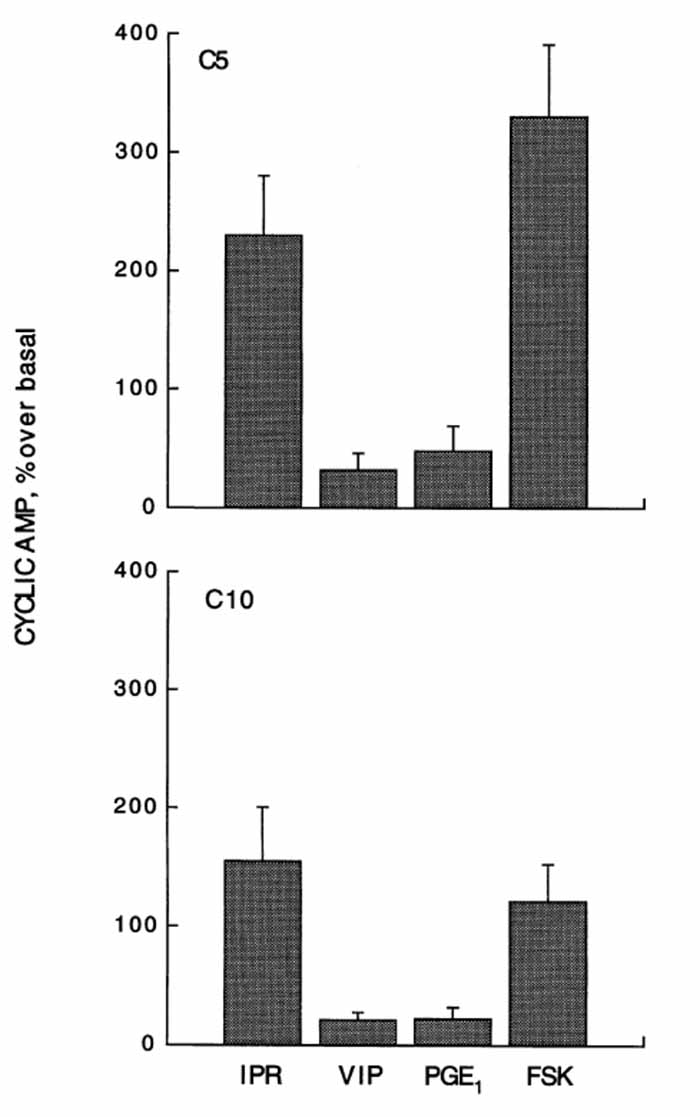
▲图8.促泌剂对3':5'-环单磷酸腺苷(环AMP)产生的影响

图9.大鼠腮腺尖细胞系C5和C10的生长情况
长期培养过程中建立了几种大鼠腮腺细胞系。遗憾的是,进一步的研究表明,这些细胞系中的大多数要么来源于导管细胞,要么是分化不良的acinarcells。为了选择和获得永生化后能充分表达其表型潜能的细胞系,需要在了解细胞特异性培养条件。直到最近,人们才确定了在原代培养中维持分化良好的大鼠唾液腺尖腺细胞的细胞培养条件:(a)使用35%0,以最大限度地表达腮腺尖腺细胞生长和分化的表型;(b)在培养基中加入地塞米松EGF和视黄酸;(c)将层粘蛋白作为细胞外基质的重要组成部分。所有这些都是为了在转染后最大限度地获得分化良好的细胞系。在开发细胞系之前,已建立了最佳的细胞生长条件和最佳的转染条件。大鼠唾液腺刺痛细胞在塑料上生长不良,也不能在培养物中继续繁殖,因此研究人员采用了选择筛选程序,而不是使用抗生素筛选。相反,在短时间后,它们会开始分化并死亡(Quissell等人,1994年)。这一事实使研究人员能够在转染后将细胞保存在细胞外基质(丝裂霉素C处理过的3T3成纤维细胞)上数个传代,使被转染的细胞在塑料上生长之前有更多的时间适应。免疫荧光抗体探针的使用和呋喃-2/AM程序对受体耦合细胞内钙动员的评估,结合光镜和电子显微镜的评估和分析,极大地促进了筛选过程。
利用pSVori-质粒是建立永生化大鼠腮腺尖突细胞的有效方法。研究人员获得了大量永生化细胞系。然而,研究人员的任务是能够快速识别和筛选出少数高度分化的细胞系。建立选择标准,利用细胞功能的多个要素对每个细胞系进行评估,极大地促进了选择过程。
虽然这些研究的目的不是为了了解细胞的永生化过程,但大T抗原的持续表达将这些细胞归入了Gluzman等人(1980年)定义的突变体的第一类或分类:即永生化细胞继续表达可检测到的大T抗原水平。腮腺细胞系C5和Cl0现已经历了30多次传代,但仍保持着中高水平的腮腺分化和功能。P物质受体介导的钙动员反应消失,而Pzu-嘌呤受体反应在初次培养和转染后显著增强,其原因尚不清楚。虽然期望永生化细胞系完全维持母细胞的正常体内表型是不现实的,但目前还不清楚这两个事件是相互关联还是巧合。要想更全面地了解这些观察结果,还需要做进一步的研究。此外,两个细胞系似乎都不表达a-淀粉酶,因为没有通过免疫荧光检测到。但是,缺乏a-淀粉酶表达并不完全是抑制作用,因为细胞在霍乱毒素存在的情况下(转染前和转染期间)受到了生长刺激。细胞内环状AMP水平的升高会降低a-淀粉酶的表达。研究发现,胆碱能和肾上腺素能受体介导的反应的维持与克隆细胞系的细胞分化程度密切相关,而肾上腺素能偶联环AMP反应则存在于所有测试的上皮细胞克隆中,与分化程度无关。需要进一步研究来确定环AMP信号转导途径是否是细胞永生化的必要条件,或者研究人员的观察结果是否只是巧合。β-肾上腺素能原位刺激会导致唾液腺尖突细胞生长急剧增加。
形态学评估表明,这两种细胞系具有中高程度的细胞分化。在研究人员的钙动员研究中,许多对胆碱能和/或肾上腺素能受体激动剂无反应的低分化细胞克隆的细胞分化程度也很低。通常情况下,在这些低分化细胞中,缺乏正常分泌功能所必需的细胞内结构,包括内质网、高尔基体、分泌颗粒和事件连接。
C5和C10细胞系都显示存在发育完全的连接复合体和末端网状结构,这很重要,因为这些形态结构对有效分泌很重要。当这两种细胞系在涂有胶原蛋白的可渗透支撑物上生长时,发现跨细胞再阻力增加。用Evomvoltohmmeter测量,C5单层细胞的跨细胞电阻为724+91欧姆/平方厘米(n=6),而C0细胞的跨细胞电阻值为75252欧姆/平方厘米(n=7)。这些数值与其他成功用于乌星室研究的上皮细胞系相比毫不逊色。研究人员还不知道其他唾液腺细胞系成功用于乌星室研究的报道。初步数据表明,ClO形成紧密的极化单层,盐离子在单层上定向移动,这种离子移动受核苷酸和毒蕈碱受体激动剂的调节。
另一方面,分泌物和其他唾液物质分泌所需的结构分布不均。有相当数量的粗面内质网,但其中大部分似乎并不活跃。高尔基体稀少,存在分泌颗粒,但数量不多,只有少数位于外泌中间。细胞大小和双相亚结构与成熟的大鼠腮腺腺细胞一致、因此,这两种细胞系都可以被描述为由中度到高度分化的腮腺尖状腺细胞组成,其中C5细胞系的分化功能最强。
要更全面地评估这两种细胞系的细胞和分泌功能水平,还需要进一步的研究。不过,安博士实验室最近的研究(Lin等人,1996年)表明,细胞系C5保留了组织特异性富脯氨酸蛋白R15表达所必需的近端和远端调控元件的基本调控蛋白。该细胞系使这些研究人员得以确定协调控制大鼠腮腺针叶富脯氨酸蛋白的组织特异性和诱导性基因表达的调控网络。这些数据表明,经过一年的培养,这两种细胞系维持了相当高的细胞分化和功能水平,因此,它们很有希望为体外研究大鼠腮腺吖丙腺细胞的分子和细胞生物学提供合适的细胞培养系统。
常见细胞污染类型如何辨别及预防解决方法:细胞培养中常见的生物污染类型有7种,分别是细菌污染,支原体污染,原虫污染,黑胶虫污染,真菌污染,病毒污染以及非细胞污染,真菌污染来源,一般是来自实验服,并且具有气候性,多雨······
细胞聚团的原因分析及如何避免:培养物中细胞可能聚集的一些原因包括:1.过度消化、2.环境压力、3.组织分解、4.过度生长、5.污染等;如何避免聚团细胞的生成;首先确认当前细胞生长密度及状态,80%左右的生长密度即可进行······
细胞有空泡原因分析及解决方法:出现细胞空泡情况有1.细胞老化2.培养液错误配制;3.细胞消化时操作不当;4.污染等等,如细胞老化,解决方法,原代细胞使用较低代次进行实验,传代细胞避免传代次数过高···
细胞半换液和全换液操作步骤:第一种方式:细胞全换液;如果是贴壁细胞,可以用全量换液法,直接吸去全部旧培养基,补充足量新鲜完全培养基;第二种方式:细胞半换液;"细胞半换液"又称"细胞半量换液",即弃掉一半旧的培养基,再······
细胞生长缓慢的可能原因有哪些:细胞培养外部因素包括细胞培养基的配方和质量问题,培养条件不理想,污染问题,细胞自身因素包含细胞的健康状态,细胞密度过高或过低,细胞老化现象,细胞特性,当细胞生长出现缓慢的问题时,我······
常用胰腺癌细胞株动物模型及胰腺癌细胞株有哪些:胰腺癌研究中常用的动物模型主要包括化学物质诱导胰腺癌动物模型,基因工程胰腺癌小鼠模型和胰腺癌移植模型,常用的胰腺细胞株MIA-PACA-2人胰腺癌细胞,Capan-2人胰腺癌细······
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
下一篇:海拉细胞的回溯: 细胞系的错误鉴定如何误导了科学文献
厦门爱恪信生物科技有限公司
手机:15859239971
邮箱:2205839769@qq.com
地址:厦门翔安火炬高新区翔星路96号建业楼D座602

微信公众号
ATCC细胞培养

技术支持
15859239971
Copyright©厦门爱恪信 闽ICP备19027235号-7
公安备案: XML地图
XML地图




 客服QQ
客服QQ