常见细胞污染类型如何辨别及预防解决方法
常见细胞污染类型如何辨别及预防解决方法:细胞培养中常见的生物污染类型有7种,分别是细菌污染,支原体污染,原虫污染,黑胶虫污染,真菌污染,病毒污染以及非细胞污染,真菌污染来源,一般是来自实验服,并且具有气候性,多雨······

发布时间:2026-07-05 09:00:00 细胞资源库平台 访问量:11
沙眼衣原体是全球最常见的性传播细菌病原体,慢性无症状感染可导致盆腔炎、输卵管瘢痕化及不孕,但现有研究多依赖二维转化细胞系(如 HeLa 细胞),无法再现子宫内膜极化上皮的结构复杂性及生理功能。尽管类器官技术已用于生殖系统研究,但尚未建立模拟衣原体自然感染途径的子宫内膜模型,且衣原体与宿主免疫细胞(如中性粒细胞)的动态相互作用机制仍不明确。
这篇发表在《Journal of Cell Science》上题为An endometrial organoid model of interactions between Chlamydia and epithelial and immune cells的研究,首次建立了沙眼衣原体(Chlamydia trachomatis)感染子宫内膜类器官的 3D 体外模型,通过模拟自然感染途径(顶端感染极化上皮),结合高分辨率成像与突变体筛选,系统解析了衣原体感染过程中的细胞骨架重塑、包涵体动态及免疫细胞招募机制,为探究衣原体在上生殖道的致病机制及药物筛选提供了贴近体内生理状态的实验体系。
子宫内膜类器官(EMOs)制备:分离小鼠子宫内膜腺体,接种于 Matrigel 中,添加 L-WRN 条件培养基(含 Wnt-3A、R-spondin-3、noggin)培养 7-10 天,形成极性完整、含黏液层及紧密连接的球形类器官;通过雌激素处理验证类器官激素响应性,荧光葡聚糖注射验证屏障功能完整性。
衣原体感染与表征:采用显微注射法将 GFP/mCherry 标记的沙眼衣原体(血清型 L2)或鼠衣原体(C. muridarum)注入类器官腔道(模拟自然顶端感染);通过共聚焦显微镜、活细胞延时成像观察感染周期(入侵、包涵体形成、释放),免疫荧光染色检测细胞骨架(F-actin、微管)、细胞连接(ZO-1、E - 钙粘蛋白)及细胞器(高尔基体)的重塑。
突变体功能验证:使用衣原体效应蛋白突变体(ΔincA、inaC缺失、cpoS缺失、Δcpaf、ΔtepP)感染类器官,检测包涵体融合、高尔基体招募、细胞死亡及免疫细胞招募等表型,明确效应蛋白的生物学功能。
免疫细胞共培养:分离小鼠骨髓来源中性粒细胞,与感染类器官共培养,通过活细胞成像量化中性粒细胞招募效率,观察其与感染上皮及包涵体的相互作用,分析cpaf和tepP突变体对中性粒细胞活化(脱颗粒、NETosis)的影响。
分子机制分析:通过免疫荧光染色检测衣原体效应蛋白与宿主蛋白的共定位,Western blot 验证效应蛋白表达,ImageJ 量化细胞骨架重塑及免疫细胞招募指标。
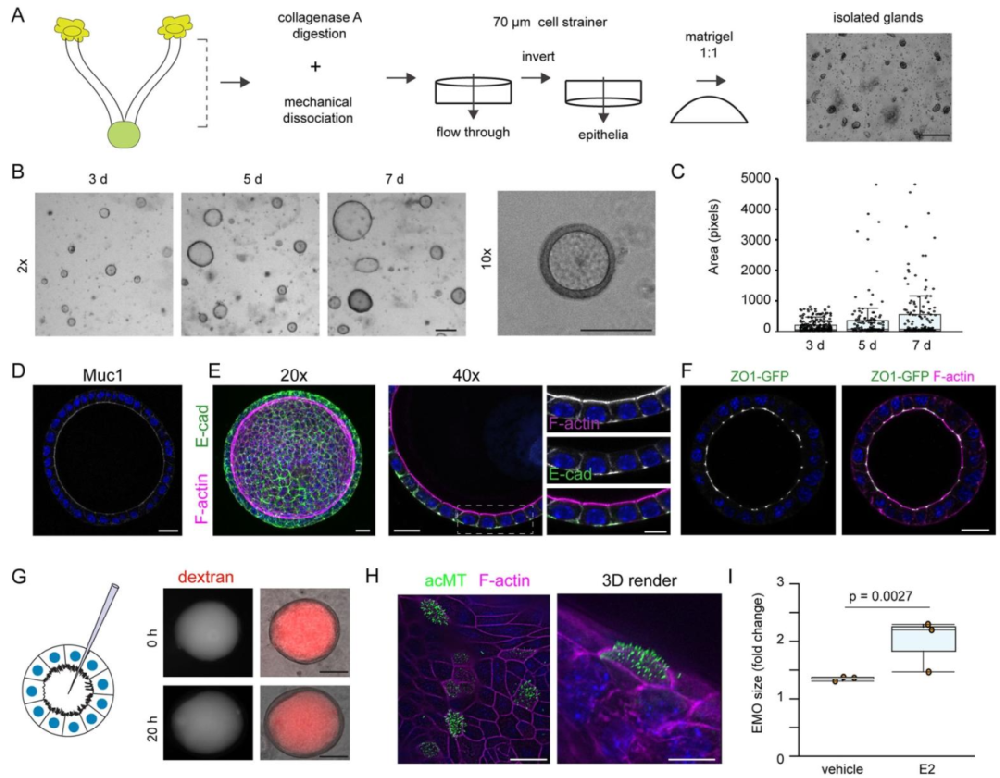
图1:子宫内膜类器官的制备与表征
该图展示类器官的制备流程及核心特性。从小鼠子宫内膜分离的腺体可高效形成球形 3D 类器官,表达黏液蛋白(MUC1),建立完整的紧密连接(ZO-1)和黏附连接(E - 钙粘蛋白),屏障功能稳定(荧光葡聚糖滞留于腔道超 20 小时);雌激素处理可促进类器官生长并诱导多纤毛上皮细胞形成,证实其激素响应性,符合子宫内膜生理特征。
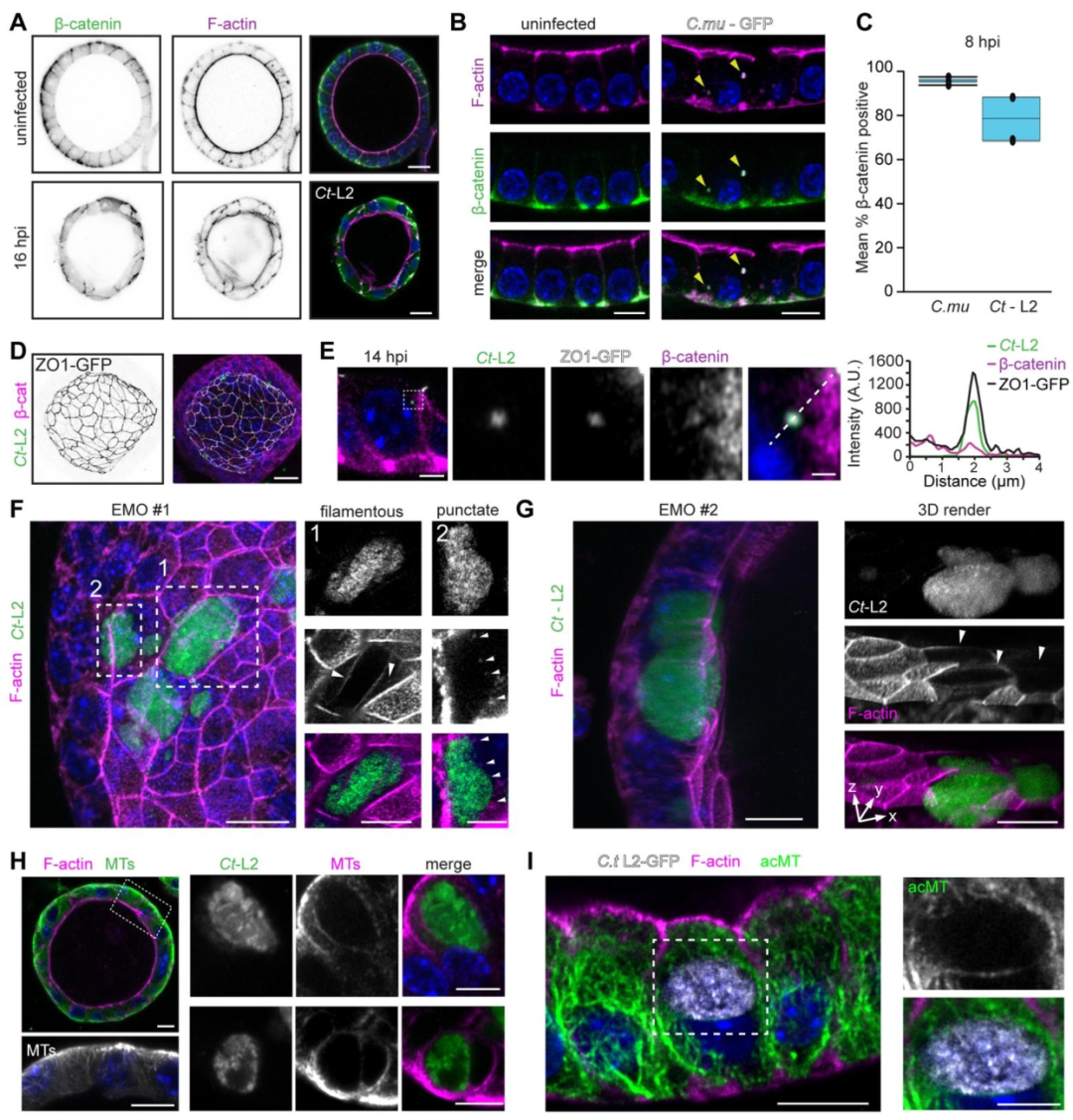
图2:衣原体感染诱导上皮细胞骨架重塑与连接破坏
衣原体通过顶端感染极化上皮后,早期即招募 β- 连环蛋白(β-catenin)和紧密连接蛋白 ZO-1 至入侵位点,导致顶端 F - 肌动蛋白(F-actin)减少及细胞连接弥散,破坏上皮极性与屏障功能;感染中期,包涵体周围形成 F - 肌动蛋白环 / 斑点、微管及乙酰化微管聚集,隔膜蛋白 2(Sept2)重组为环绕包涵体的丝状环,角蛋白也重新定位至包涵体周围,证实衣原体通过保守机制重塑宿主细胞骨架以维持包涵体稳定。
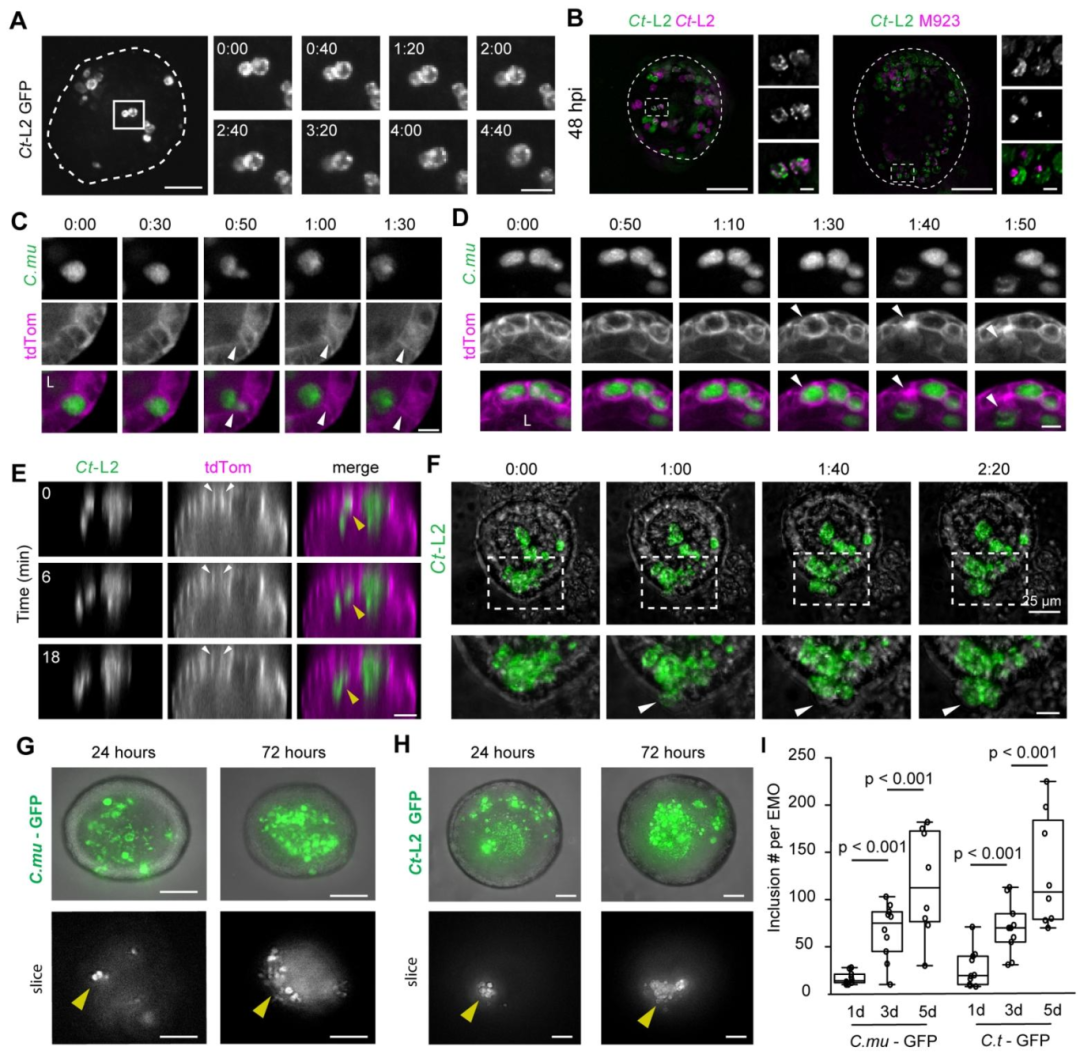
图3:包涵体动态变化与病原体传播
活细胞成像显示,衣原体包涵体可发生融合(依赖 IncA 蛋白),鼠衣原体主要通过顶端挤出 / 裂解释放至类器官腔道,而沙眼衣原体可同时通过顶端和基底侧释放,实现类器官内二次感染(感染后 5 天包涵体数量显著增加);该结果首次在 3D 极化上皮中证实衣原体释放途径的菌株特异性差异,为理解其组织扩散机制提供依据。
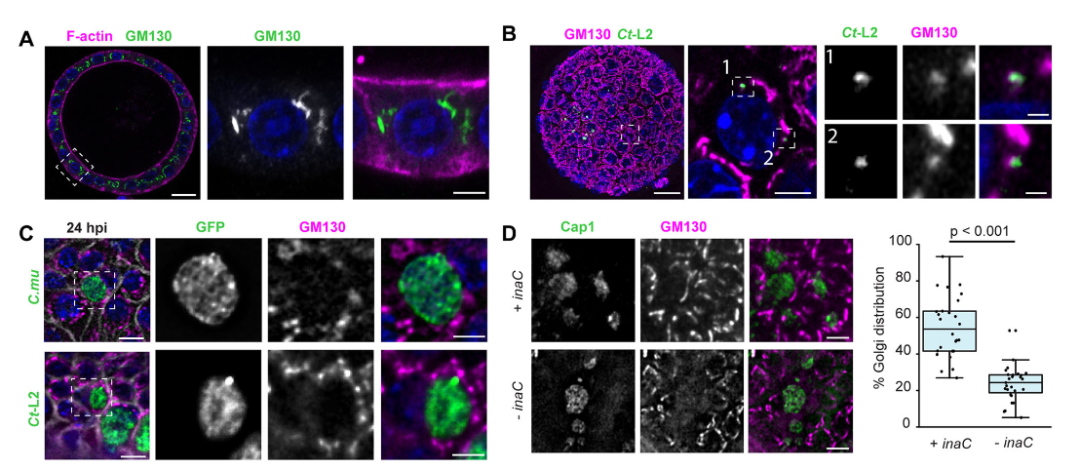
图4:效应蛋白 InaC 调控高尔基体重组
衣原体感染可诱导宿主高尔基体片段化并招募至包涵体周围,该过程依赖包涵体膜蛋白 InaC(CT813);inaC缺失突变体无法有效招募高尔基体片段,而互补菌株可恢复表型,证实 InaC 通过调控高尔基体定位为衣原体复制提供营养支持,且该机制在极化上皮中高度保守。

图5:效应蛋白 CpoS 抑制感染诱导的细胞死亡
衣原体通过效应蛋白 CpoS(CT229)抑制宿主细胞死亡,野生型感染类器官中碘化丙啶(PI)染色阳性细胞极少,而cpoS缺失突变体感染后,包涵体破裂伴随细胞肿胀、挤出及膜完整性丧失,PI 阳性细胞及包涵体比例显著升高,证实 CpoS 通过维持包涵体完整性阻断细胞自主免疫防御中的细胞死亡通路。
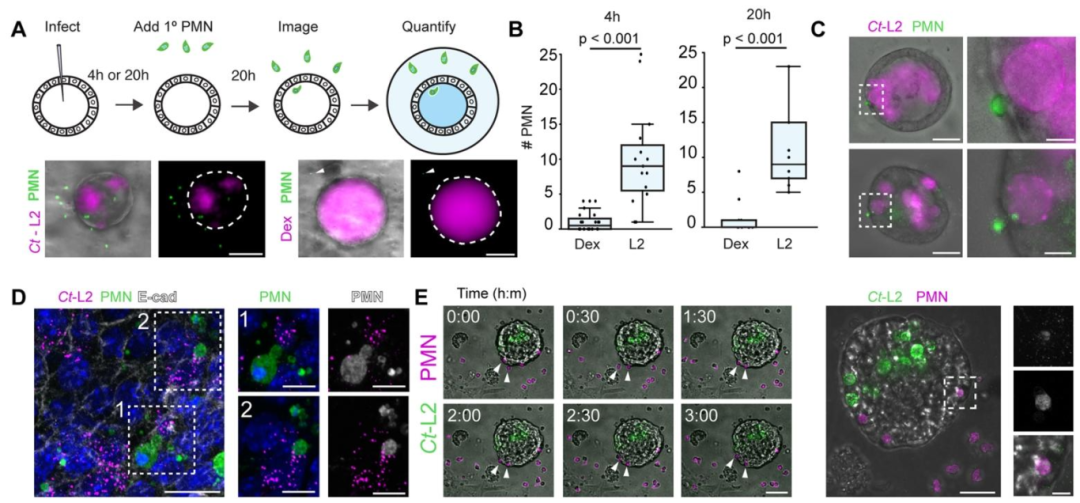
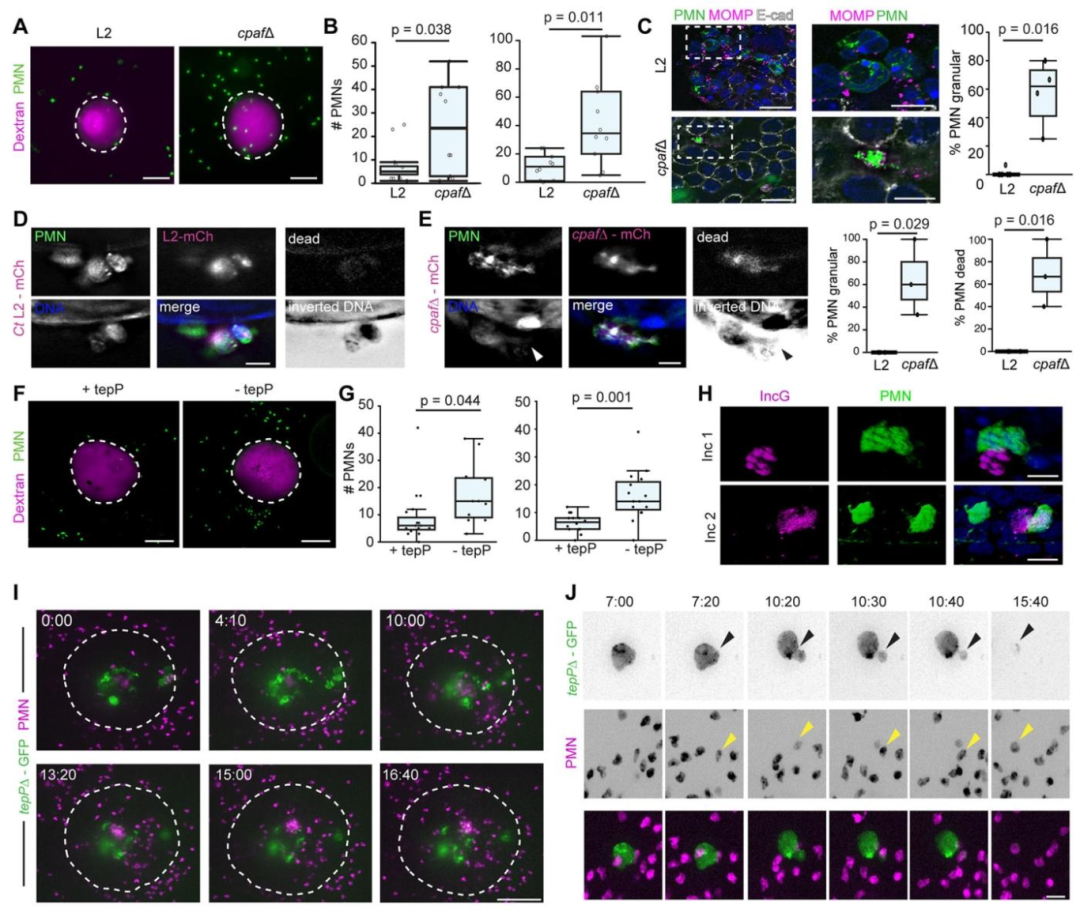
图6-7:衣原体调控中性粒细胞招募与活化
中性粒细胞可特异性招募至感染类器官(非感染类器官无显著招募),并直接接触感染上皮及包涵体;衣原体效应蛋白 CPAF(CT858)通过切割中性粒细胞表面受体抑制其招募及 NETosis(中性粒细胞胞外诱捕网形成),而 Δcpaf突变体感染后中性粒细胞招募增加且出现颗粒化表型及膜完整性丧失;效应蛋白 TepP(CT875)通过抑制趋化因子(如 IL-6、CXCL3)表达减少中性粒细胞招募,ΔtepP突变体感染可诱导中性粒细胞聚集、腔道浸润及包涵体吞噬,揭示衣原体通过多重效应蛋白调控先天免疫应答以逃逸清除。
本研究建立了首个模拟衣原体自然感染途径的子宫内膜类器官模型,证实该模型可再现衣原体感染的关键过程(入侵、包涵体形成、释放及二次感染),且衣原体在极化上皮中采用与二维细胞系保守的宿主调控机制(细胞骨架重塑、细胞器招募)。核心发现包括:1)衣原体通过 InaC 调控高尔基体重组、CpoS 抑制细胞死亡维持复制微环境;2)CPAF 和 TepP 通过不同机制抑制中性粒细胞招募与活化,参与免疫逃逸;3)鼠衣原体与沙眼衣原体存在释放途径差异,可能与组织扩散能力相关。
该模型的优势在于保留子宫内膜上皮极性、激素响应性及屏障功能,可结合突变体筛选与免疫细胞共培养,解析病原体 - 上皮 - 免疫细胞的三维相互作用。局限性包括未涵盖子宫内膜间质细胞,且未探究激素对感染的调控作用。未来可通过构建上皮 - 间质类器官组装体、结合人类子宫内膜类器官及临床菌株,进一步验证致病机制并开发靶向治疗策略。
常见细胞污染类型如何辨别及预防解决方法:细胞培养中常见的生物污染类型有7种,分别是细菌污染,支原体污染,原虫污染,黑胶虫污染,真菌污染,病毒污染以及非细胞污染,真菌污染来源,一般是来自实验服,并且具有气候性,多雨······
细胞聚团的原因分析及如何避免:培养物中细胞可能聚集的一些原因包括:1.过度消化、2.环境压力、3.组织分解、4.过度生长、5.污染等;如何避免聚团细胞的生成;首先确认当前细胞生长密度及状态,80%左右的生长密度即可进行······
细胞有空泡原因分析及解决方法:出现细胞空泡情况有1.细胞老化2.培养液错误配制;3.细胞消化时操作不当;4.污染等等,如细胞老化,解决方法,原代细胞使用较低代次进行实验,传代细胞避免传代次数过高···
细胞半换液和全换液操作步骤:第一种方式:细胞全换液;如果是贴壁细胞,可以用全量换液法,直接吸去全部旧培养基,补充足量新鲜完全培养基;第二种方式:细胞半换液;"细胞半换液"又称"细胞半量换液",即弃掉一半旧的培养基,再······
细胞生长缓慢的可能原因有哪些:细胞培养外部因素包括细胞培养基的配方和质量问题,培养条件不理想,污染问题,细胞自身因素包含细胞的健康状态,细胞密度过高或过低,细胞老化现象,细胞特性,当细胞生长出现缓慢的问题时,我······
常用胰腺癌细胞株动物模型及胰腺癌细胞株有哪些:胰腺癌研究中常用的动物模型主要包括化学物质诱导胰腺癌动物模型,基因工程胰腺癌小鼠模型和胰腺癌移植模型,常用的胰腺细胞株MIA-PACA-2人胰腺癌细胞,Capan-2人胰腺癌细······
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
上一篇:突破黑色素瘤脑转移治疗瓶颈:皮质类器官模型助力精准药物筛选
下一篇:突破实体瘤免疫治疗瓶颈:类器官 + 外周血淋巴细胞共培养,高效诱导肿瘤反应性 T 细胞
厦门爱恪信生物科技有限公司
手机:15859239971
邮箱:2205839769@qq.com
地址:厦门翔安火炬高新区翔星路96号建业楼D座602

微信公众号
ATCC细胞培养

技术支持
15859239971
Copyright©厦门爱恪信 闽ICP备19027235号-7
公安备案: XML地图
XML地图




 客服QQ
客服QQ