常见细胞污染类型如何辨别及预防解决方法
常见细胞污染类型如何辨别及预防解决方法:细胞培养中常见的生物污染类型有7种,分别是细菌污染,支原体污染,原虫污染,黑胶虫污染,真菌污染,病毒污染以及非细胞污染,真菌污染来源,一般是来自实验服,并且具有气候性,多雨······

发布时间:2026-07-05 17:00:00 细胞资源库平台 访问量:7
mRNA 疫苗凭借快速开发、高免疫原性和良好安全性,已成为癌症免疫治疗的核心方向之一,个性化 mRNA 疫苗联合免疫检查点抑制剂在黑色素瘤、胰腺癌等癌症中取得了积极的临床结果。但现有 mRNA 疫苗平台存在明显局限:传统线性 mRNA 需 5’加帽和核苷酸修饰才能保证稳定性和翻译效率,增加了生产复杂度;环状 RNA(circRNA)生产流程繁琐、成本高昂;自扩增 mRNA(saRNA)易产生双链 RNA(dsRNA),引发系统性炎症反应。此外,多数 mRNA 疫苗缺乏内在佐剂活性,需额外添加佐剂才能激活足够的免疫响应。
由复旦大学附属肿瘤医院、福建医科大学附属肿瘤医院的团队发表在《Nature Communications》的研究:An engineered linear cap-independent mRNA vaccine with intrinsic adjuvanticity induces potent anti-tumor immunity in mice,团队通过融合病毒来源的外切核酸酶抗性元件与 RNA 结合蛋白 motif,构建了无需 5’加帽和核苷酸修饰的线性帽非依赖 mRNA(LciRNA)疫苗平台,该疫苗兼具高稳定性、高效翻译效率和内在佐剂活性,在小鼠肿瘤模型中实现了强效抗肿瘤免疫响应。
1.核心元件筛选与优化:从 5 种黄病毒(登革热病毒、寨卡病毒等)的亚基因组 RNA(sfRNA)中筛选外切核酸酶抗性元件(xrRNA),通过荧光素酶报告基因筛选出最优的 UX1 元件(来源于 Usutu 病毒);在 UX1 下游融合多聚腺苷酸结合蛋白(PABP)结合基序,形成 UPA 保护序列;搭配优化的肠道病毒 A(EV-A)内部核糖体进入位点(IRES),构建基础 LciRNA 框架;进一步筛选人类 β- 珠蛋白来源的 5’和 3’非翻译区(UTR),优化翻译效率。
2.LciRNA 疫苗构建与制备:将目标抗原基因(OVA、HPV E6/E7)插入 LciRNA 框架,通过体外转录合成 LciRNA(无需添加帽类似物或修饰核苷酸);同时制备传统加帽 mRNA(CAP)和 m1Ψ 修饰的加帽 mRNA(mCAP)作为对照;将各类 mRNA 封装于 SM102 脂质纳米颗粒(LNP)中,用于体内外实验。
3.体外功能与机制验证:通过荧光素酶报告基因实验验证 LciRNA 的稳定性和翻译效率;利用 XRN-1 外切核酸酶消化实验明确 UPA 序列的抗降解作用;通过 RNA pull-down、质谱分析和 RNA 免疫沉淀(RIP)鉴定与 UPA 结合的 RNA 结合蛋白(RBP);刺激小鼠骨髓来源树突状细胞(BMDC),通过 qRT-PCR 和 RNA-seq 分析其免疫激活能力。
4.体内疗效与安全性评估:在 B16F10-OVA 黑色素瘤和 HPV 相关 TC-1 肿瘤小鼠模型中,评估 LciRNA 疫苗的抗肿瘤效果;通过流式细胞术分析外周血、脾脏和肿瘤组织中的抗原特异性 T 细胞响应;监测小鼠体重、血清转氨酶水平和器官组织病理学变化,评估疫苗安全性。
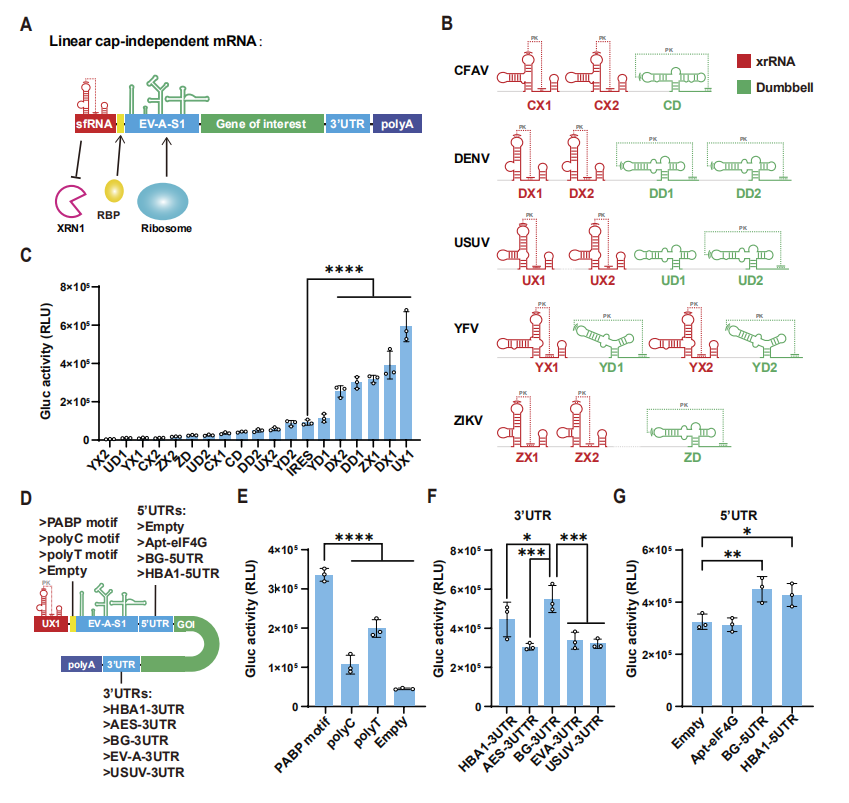
图1:LciRNA 疫苗的设计与核心元件优化
该图呈现了 LciRNA 的结构设计与关键元件筛选过程:LciRNA 的 5’端融合黄病毒 sfRNA 来源的 xrRNA 元件和 PABP 结合基序(UPA 序列),中间为 EV-A-S1 IRES,下游为目标抗原基因和 3’UTR 及 poly (A) 尾,无需 5’加帽即可启动翻译。通过荧光素酶报告基因筛选,从 18 种候选 xrRNA 元件中鉴定出 UX1(Usutu 病毒来源)具有最强的表达增强效果;进一步优化发现,在 UX1 和 IRES 之间插入 PABP 基序,搭配人类 β- 珠蛋白的 5’和 3’UTR,可显著提升 LciRNA 的翻译效率,为后续疫苗构建奠定了基础。
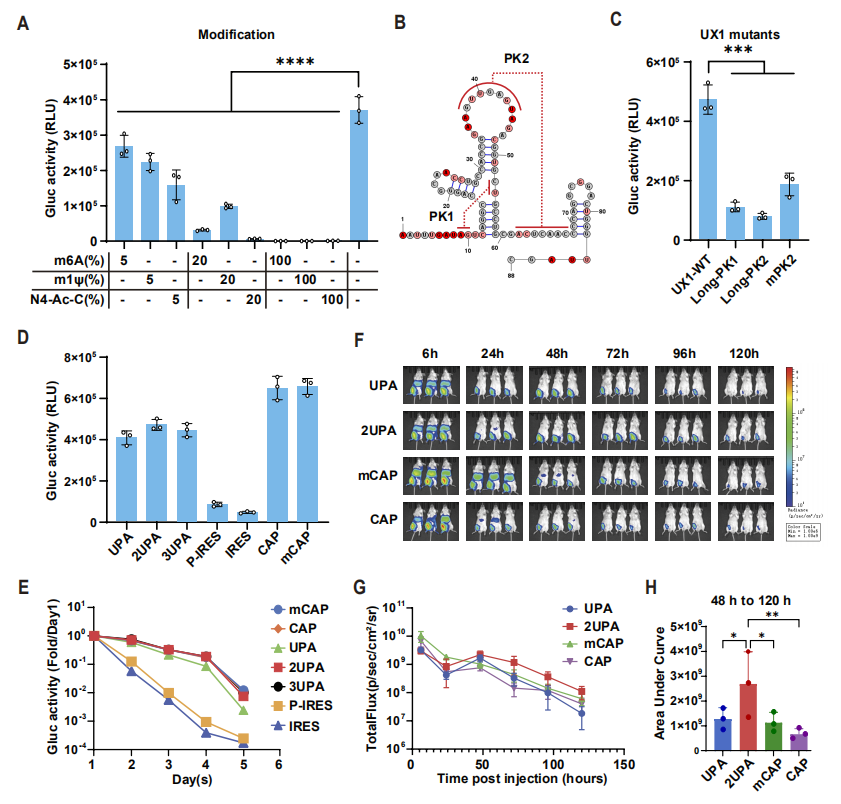
图2:LciRNA 的稳定性与体内表达优势
该图验证了 LciRNA 的稳定性及与传统 mRNA 的表达差异:核苷酸修饰(m6A、m1Ψ 等)会破坏 UX1 和 IRES 的结构,导致 LciRNA 表达下降,表明 LciRNA 无需修饰即可高效表达;对 UX1 的伪结结构进行突变后,其抗降解能力显著减弱,证实天然 UX1 序列已达到最优抗降解效果。体内外稳定性实验显示,含双 UPA 序列(2UPA)的 LciRNA 在细胞中可维持 5 天稳定表达,与加帽 mRNA 相当;在小鼠体内,2UPA-LciRNA 的生物发光信号在 48 小时后超过 m1Ψ 修饰的加帽 mRNA(mCAP),且持续至 120 小时,表明其体内稳定性更优,为长效抗原表达和免疫激活提供了保障。
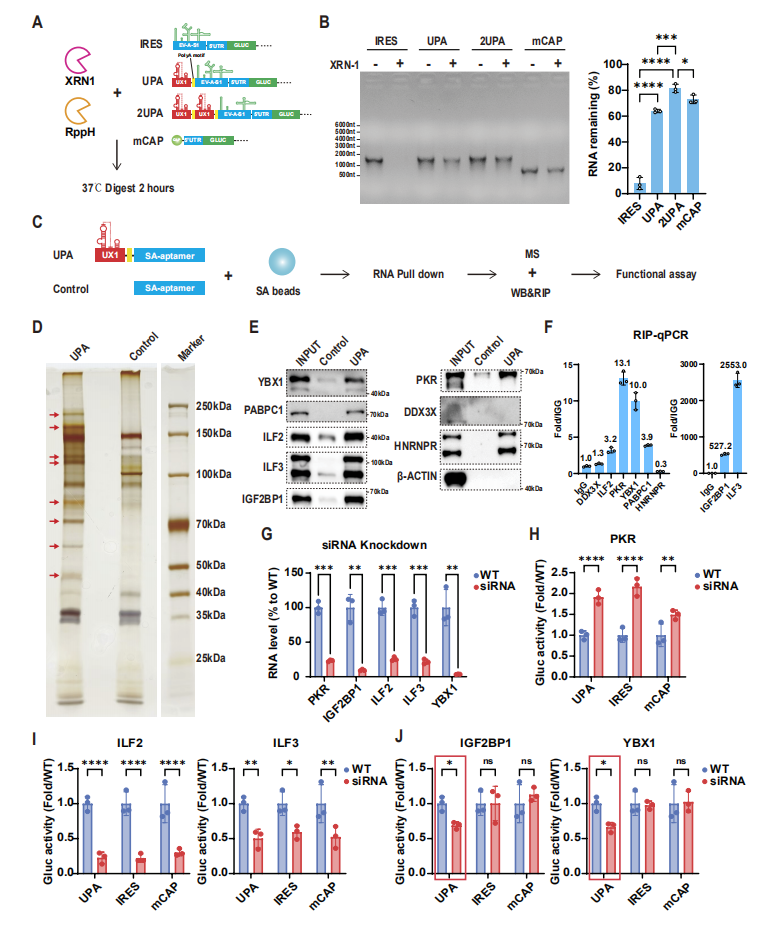
图3:UPA 序列通过抑制 XRN-1 降解和结合 RBP 增强 LciRNA 稳定性
该图揭示了 LciRNA 的稳定性机制:体外 XRN-1 消化实验显示,未受保护的 RNA 在 2 小时内完全降解,而 UPA-LciRNA 和 2UPA-LciRNA 与加帽 mRNA 类似,能有效抵抗 XRN-1 介导的外切核酸酶降解,且 2UPA 的保护效果优于单 UPA。通过 RNA pull-down 结合质谱分析,鉴定出 6 种与 UPA 序列特异性结合的 RNA 结合蛋白(RBP),包括 PABPC1、IGF2BP1、YBX1 等已知的 mRNA 稳定蛋白;siRNA 敲低实验证实,敲低 IGF2BP1 或 YBX1 会特异性降低 LciRNA 的表达,而对加帽 mRNA 无影响,表明 UPA 序列通过 “抑制 XRN-1 降解 + 招募稳定型 RBP” 双重机制,实现了无需加帽的高稳定性。
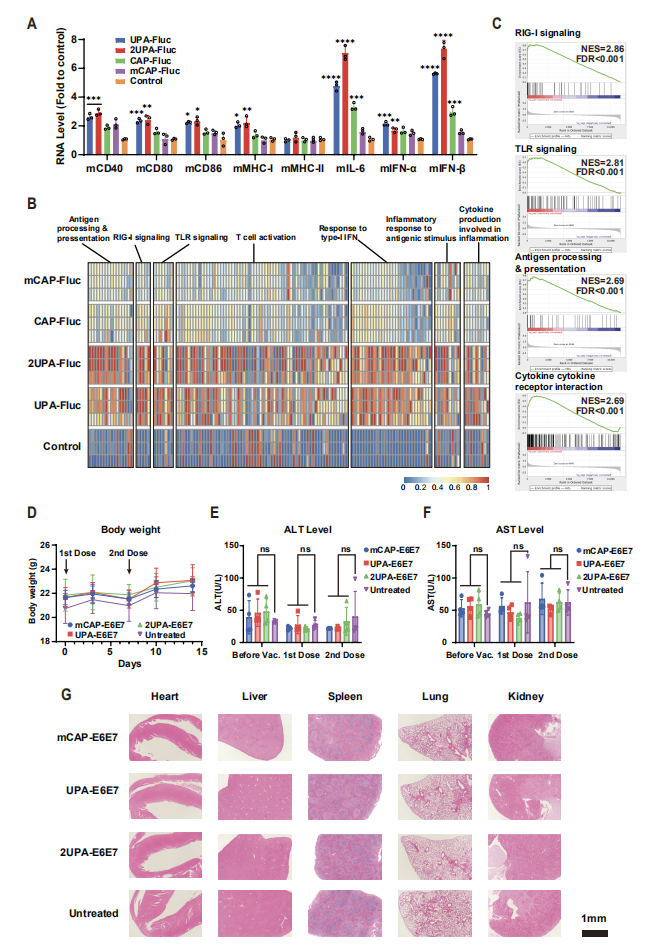
图4:LciRNA 具有内在佐剂活性,可激活 DC 成熟且安全性良好
该图展示了 LciRNA 的免疫激活能力与安全性:刺激小鼠 BMDC 后,LciRNA(UPA-Fluc、2UPA-Fluc)能显著上调 MHC-I、CD40、CD80 等共刺激分子的表达,以及 IFN-α、IFN-β、IL-6 等促炎细胞因子的分泌,其激活效果显著强于传统加帽 mRNA(CAP-Fluc、mCAP-Fluc);RNA-seq 分析显示,LciRNA 强烈激活了模式识别受体(PRR)信号通路、抗原加工呈递通路和 I 型干扰素响应通路,证实其通过多 PRR engagement 发挥内在佐剂作用。安全性评估显示,LciRNA 疫苗接种后,小鼠体重无明显变化,血清转氨酶(ALT、AST)水平与对照组相当,主要器官无病理损伤,表明其在有效剂量下具有良好的生物安全性。
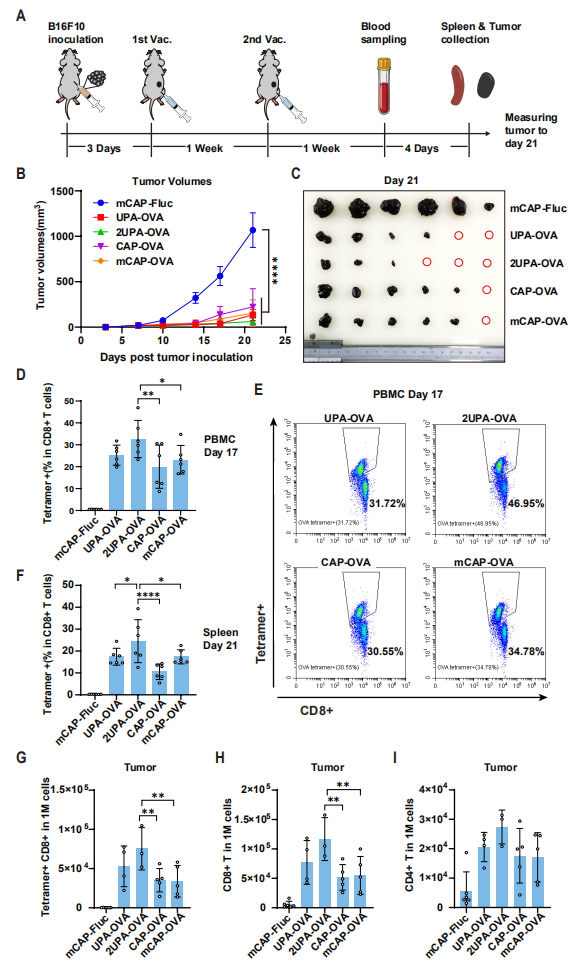
图5:LciRNA 疫苗在黑色素瘤模型中诱导强效抗肿瘤 T 细胞响应
该图验证了 LciRNA 疫苗在 B16F10-OVA 黑色素瘤模型中的治疗效果:接种 2UPA-OVA LciRNA 疫苗的小鼠,肿瘤生长受到显著抑制,21 天内 3/6 的小鼠实现完全肿瘤消退,疗效优于传统加帽 mRNA 疫苗。流式细胞术分析显示,2UPA-OVA 疫苗诱导的 OVA 特异性 CD8+ T 细胞比例(32.6%)显著高于 UPA-OVA(25.3%)和 mCAP-OVA(23.2%);肿瘤浸润淋巴细胞分析显示,LciRNA 疫苗组的 CD4 + 和 CD8+ T 细胞浸润数量显著增加,且 OVA 特异性 CD8+ T 细胞比例更高,表明 LciRNA 疫苗能有效将 “冷肿瘤” 转化为 “热肿瘤”,增强肿瘤局部免疫响应。
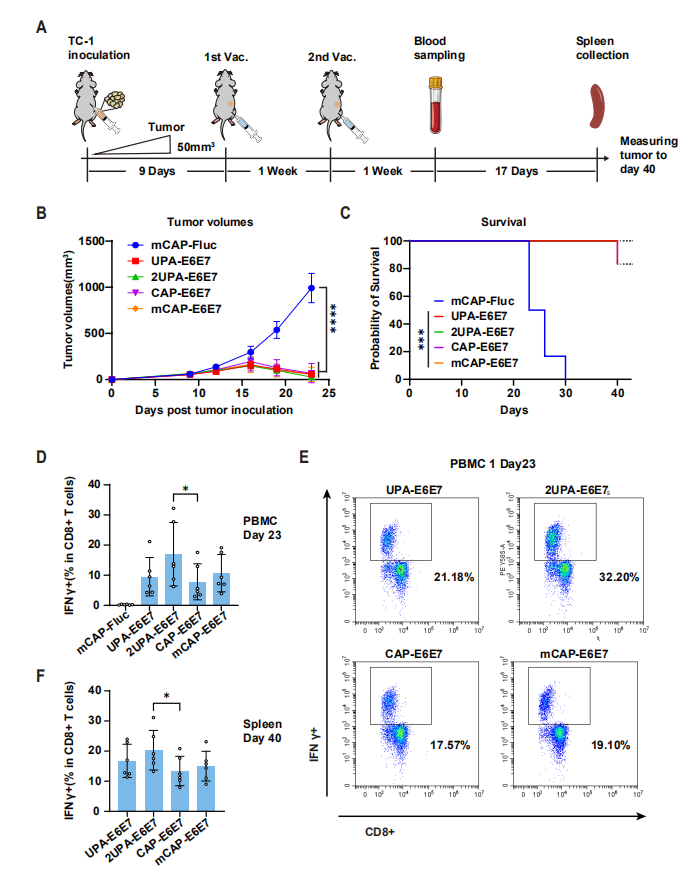
图6:LciRNA 疫苗有效清除 HPV 相关肿瘤并诱导长效免疫记忆
该图展示了 LciRNA 疫苗在 HPV 相关肿瘤模型中的治疗效果:针对 HPV E6/E7 抗原的 2UPA-E6E7 LciRNA 疫苗,在 TC-1 肿瘤小鼠模型中实现了持续的肿瘤消退,所有接种小鼠在 40 天观察期内均无复发,而传统加帽 mRNA 疫苗组有部分小鼠出现肿瘤复发。免疫分析显示,2UPA-E6E7 疫苗诱导的 E6/E7 特异性 IFN-γ+ CD8+ T 细胞比例(17.0%)显著高于加帽 mRNA 组;40 天后脾脏中仍能检测到高比例的抗原特异性 CD8+ T 细胞,表明 LciRNA 疫苗能诱导长效肿瘤特异性免疫记忆,为预防肿瘤复发提供了保障。
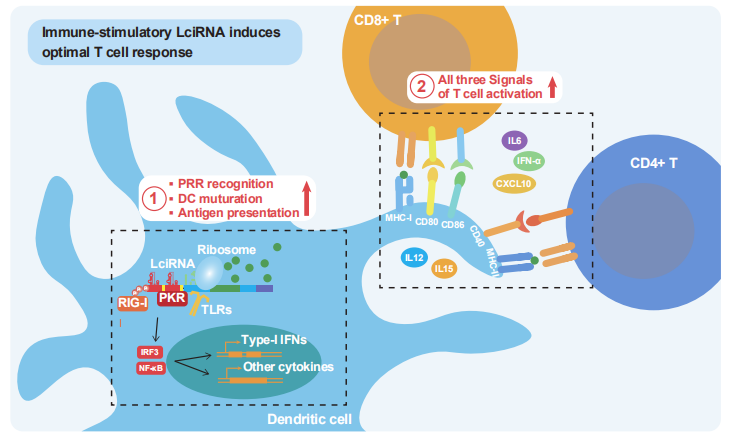
图7:LciRNA 疫苗激活抗肿瘤免疫的分子机制示意图
该图总结了 LciRNA 疫苗的作用机制:LciRNA 被树突状细胞(DC)摄取后,其 5’三磷酸基团、病毒来源的 IRES 结构及 UPA 序列分别被 RIG-I、PKR、TLR 等模式识别受体识别,激活 NF-κB 和 IRF3 信号通路,促进 I 型干扰素和促炎细胞因子(IL-6、IL-12 等)分泌;同时,LciRNA 的高稳定性保证了抗原的持续表达和呈递,DC 表面共刺激分子(CD40、CD80、CD86)表达上调;最终,DC 通过 “抗原呈递 + 共刺激信号 + 细胞因子” 三重信号,高效激活肿瘤特异性 CD4 + 和 CD8+ T 细胞,增强外周免疫响应和肿瘤浸润,实现肿瘤清除和长期免疫记忆。
本研究通过序列工程构建了无需 5’加帽和核苷酸修饰的 LciRNA 疫苗平台,该平台融合黄病毒来源的 UX1 外切核酸酶抗性元件与 PABP 结合基序,形成 UPA 保护序列,搭配优化的 EV-A IRES,实现了高效翻译和高稳定性;同时,LciRNA 通过多模式识别受体激活,具有内在佐剂活性,可促进 DC 成熟和促炎因子分泌,无需额外添加佐剂。在小鼠黑色素瘤和 HPV 相关肿瘤模型中,LciRNA 疫苗展现出优于传统加帽 mRNA 疫苗的抗肿瘤效果,能诱导强效、长效的肿瘤特异性 T 细胞响应和免疫记忆,且安全性良好。该研究解决了现有 mRNA 疫苗平台 “疗效依赖复杂修饰或佐剂” 的核心痛点,提供了一种低成本、高疗效、易生产的癌症疫苗设计方案,为 mRNA 疫苗的临床转化和广泛应用奠定了基础。未来需进一步开展大动物实验和临床试验,验证 LciRNA 疫苗的长期安全性和临床疗效,同时优化元件组合以提升其在不同癌症类型中的适用性。
常见细胞污染类型如何辨别及预防解决方法:细胞培养中常见的生物污染类型有7种,分别是细菌污染,支原体污染,原虫污染,黑胶虫污染,真菌污染,病毒污染以及非细胞污染,真菌污染来源,一般是来自实验服,并且具有气候性,多雨······
细胞聚团的原因分析及如何避免:培养物中细胞可能聚集的一些原因包括:1.过度消化、2.环境压力、3.组织分解、4.过度生长、5.污染等;如何避免聚团细胞的生成;首先确认当前细胞生长密度及状态,80%左右的生长密度即可进行······
细胞有空泡原因分析及解决方法:出现细胞空泡情况有1.细胞老化2.培养液错误配制;3.细胞消化时操作不当;4.污染等等,如细胞老化,解决方法,原代细胞使用较低代次进行实验,传代细胞避免传代次数过高···
细胞半换液和全换液操作步骤:第一种方式:细胞全换液;如果是贴壁细胞,可以用全量换液法,直接吸去全部旧培养基,补充足量新鲜完全培养基;第二种方式:细胞半换液;"细胞半换液"又称"细胞半量换液",即弃掉一半旧的培养基,再······
细胞生长缓慢的可能原因有哪些:细胞培养外部因素包括细胞培养基的配方和质量问题,培养条件不理想,污染问题,细胞自身因素包含细胞的健康状态,细胞密度过高或过低,细胞老化现象,细胞特性,当细胞生长出现缓慢的问题时,我······
常用胰腺癌细胞株动物模型及胰腺癌细胞株有哪些:胰腺癌研究中常用的动物模型主要包括化学物质诱导胰腺癌动物模型,基因工程胰腺癌小鼠模型和胰腺癌移植模型,常用的胰腺细胞株MIA-PACA-2人胰腺癌细胞,Capan-2人胰腺癌细······
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
下一篇:GLP-1R 信号激活促进高氧暴露新生大鼠肺发育:线粒体代谢与 m6A 甲基化调控机制
厦门爱恪信生物科技有限公司
手机:15859239971
邮箱:2205839769@qq.com
地址:厦门翔安火炬高新区翔星路96号建业楼D座602

微信公众号
ATCC细胞培养

技术支持
15859239971
Copyright©厦门爱恪信 闽ICP备19027235号-7
公安备案: XML地图
XML地图




 客服QQ
客服QQ