常见细胞污染类型如何辨别及预防解决方法
常见细胞污染类型如何辨别及预防解决方法:细胞培养中常见的生物污染类型有7种,分别是细菌污染,支原体污染,原虫污染,黑胶虫污染,真菌污染,病毒污染以及非细胞污染,真菌污染来源,一般是来自实验服,并且具有气候性,多雨······

发布时间:2026-04-18 17:00:05 细胞资源库平台 访问量:83
胃肠道类器官(肠类器官、结肠类器官、胃类器官)作为成人组织干细胞衍生的 3D 上皮模型,可复现肠道屏障完整性、极性分泌 / 吸收等核心生理功能,是研究上皮生物学、感染性疾病及药物筛选的关键工具。但传统类器官(如 Sato 法构建)呈 “顶 - 内(apical-in)” 极性 —— 顶膜(与肠腔接触的功能面,负责营养吸收、微生物互作)包裹于内部腔室,基底膜朝向外部 ECM,导致顶膜难以直接访问,严重限制了宿主 - 病原体互作、顶膜特异性药物测试等研究。现有解决方法(微注射、机械破碎、2D 单层培养)存在技术复杂、破坏上皮结构或失去 3D 生理相关性等缺陷,为此研究团队开发了悬浮培养介导的极性反转技术,可在 24-48 小时内将类器官转化为 “顶 - 外(apical-out)” 极性(顶膜朝向培养基),同时保留上皮完整性与功能,填补顶膜易访问 3D 模型的空白。
来自美国斯坦福大学儿科传染病科、微生物学与免疫学系的团队在《Nature Protocols》上发表题为Controlling the polarity of human gastrointestinal organoids to investigate epithelial biology and infectious diseases的实验方案研究。
顶 - 外类器官制备流程:从人 / 小鼠胃肠道组织分离隐窝,在 BME 中培养为顶 - 内类器官(7-10 天,“气泡状” 囊肿),经冰浴 5mM EDTA/PBS 解离 BME(4℃旋转孵育 1 小时),300g 离心收集后用 DMEM 洗涤,重悬于分化培养基(含 EGF、Noggin、Gastrin 等)接种至超低吸附板,24-48 小时完成极性反转,通过 F-actin(顶膜微绒毛)、ZO-1(紧密连接)、β- 连环蛋白(基底膜)免疫荧光验证极性;
核心机制:EDTA 去除 BME 后,破坏 ECM 与上皮细胞 β1 整合素的相互作用,上皮细胞发生整体形态外翻,顶膜转向外部;
应用体系:用于感染性疾病研究(沙门氏菌、李斯特菌等病原体感染机制及药物筛选)、上皮功能检测(FITC - 葡聚糖扩散实验测屏障功能、荧光脂肪酸吸收实验测营养吸收),且悬浮培养基添加 20% BME 可维持顶 - 内极性,用于对比实验。
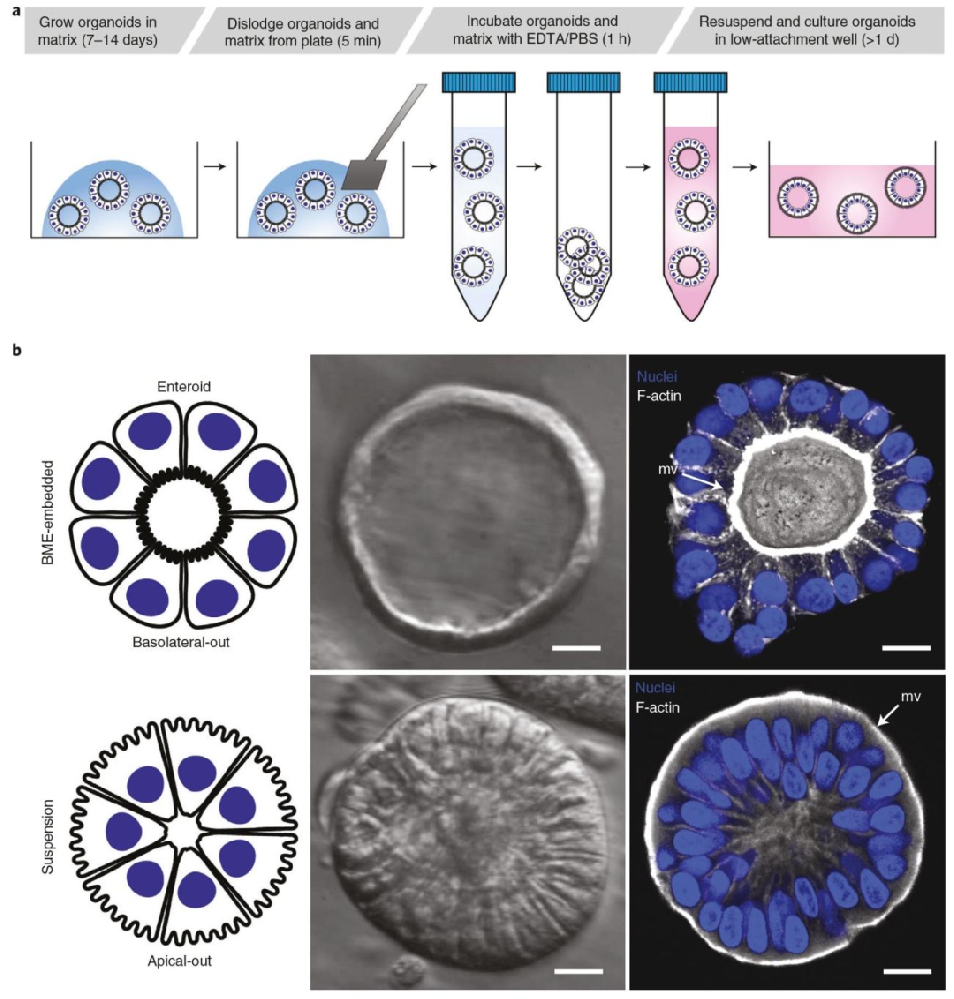
图 1:顶 - 外胃肠道类器官制备流程与极性对比
该图展示极性反转的实验设计与形态差异,(A)为流程示意图:从 BME 中顶 - 内类器官开始,经 EDTA 解离 BME、悬浮培养形成顶 - 外类器官;(B)为形态与染色对比:顶 - 内类器官(BME 中)呈 “气泡状”、内部有腔室,顶膜微绒毛(F-actin 染色,白)朝向腔室;顶 - 外类器官(悬浮中)无明显腔室,微绒毛朝向外部,共聚焦 3D 重建显示 F-actin 在顶 - 外类器官表面富集,DAPI(蓝)标记细胞核,证明 24 小时悬浮培养可实现类器官极性外翻且保留上皮结构。
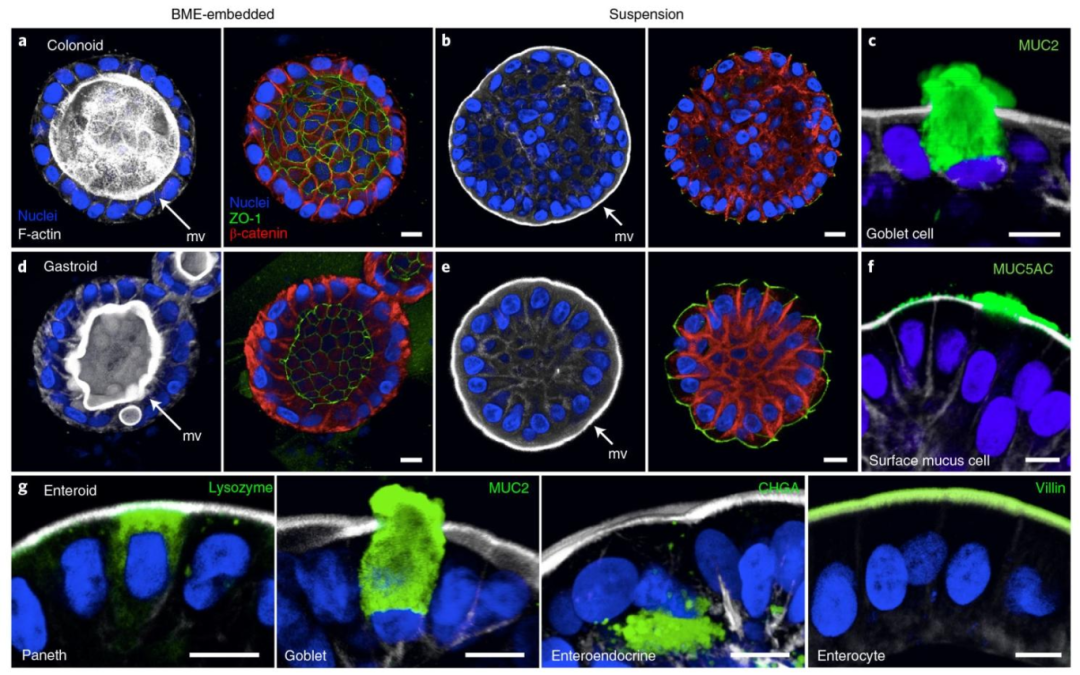
图 2:顶 - 外类器官的紧密连接与多谱系分化
该图验证顶 - 外类器官的结构完整性与分化能力,(A/B/D/E)显示结肠类器官(A/B)和胃类器官(D/E)中,顶 - 外类器官的 ZO-1(紧密连接,绿)分布于细胞表面,β- 连环蛋白(基底膜,红)位于细胞内侧,F-actin(微绒毛,白)富集于外部,与顶 - 内类器官标记分布相反;(C/F/G)显示结肠类器官表达杯状细胞标记 MUC2(绿),胃类器官表达表面黏液细胞标记 MUC5AC(绿),肠类器官表达潘氏细胞(溶菌酶)、肠内分泌细胞(嗜铬粒蛋白 A)、肠细胞(绒毛蛋白)标记,证明顶 - 外类器官保留紧密连接完整性且可正常分化为胃肠道上皮所有细胞类型。
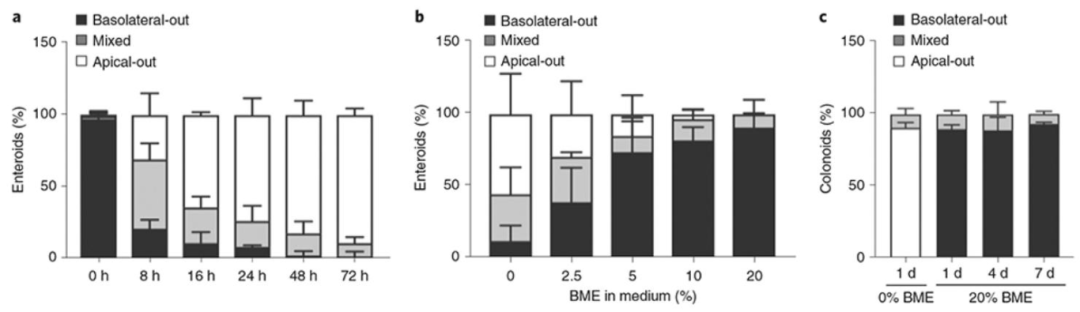
图 3:极性反转动力学与 BME 的剂量依赖性
该图量化极性反转的时间规律与 BME 的调控作用,(A)时间动力学显示悬浮培养 0-48 小时,顶 - 外类器官比例从 0% 升至 90%,24 小时达 80%(数据来自 3 次实验共 900 个类器官);(B)BME 剂量效应显示培养基中 BME 浓度 0%-20% 时,顶 - 外类器官比例从 90% 降至 5%,20% BME 完全抑制反转(数据来自 3 次实验共 303 个类器官);(C)长期稳定性显示 20% BME 培养的结肠类器官 7 天内顶 - 内极性维持率超 95%,证明极性反转具有时间依赖性,BME 可剂量依赖性抑制反转,为同批次对比实验提供条件。
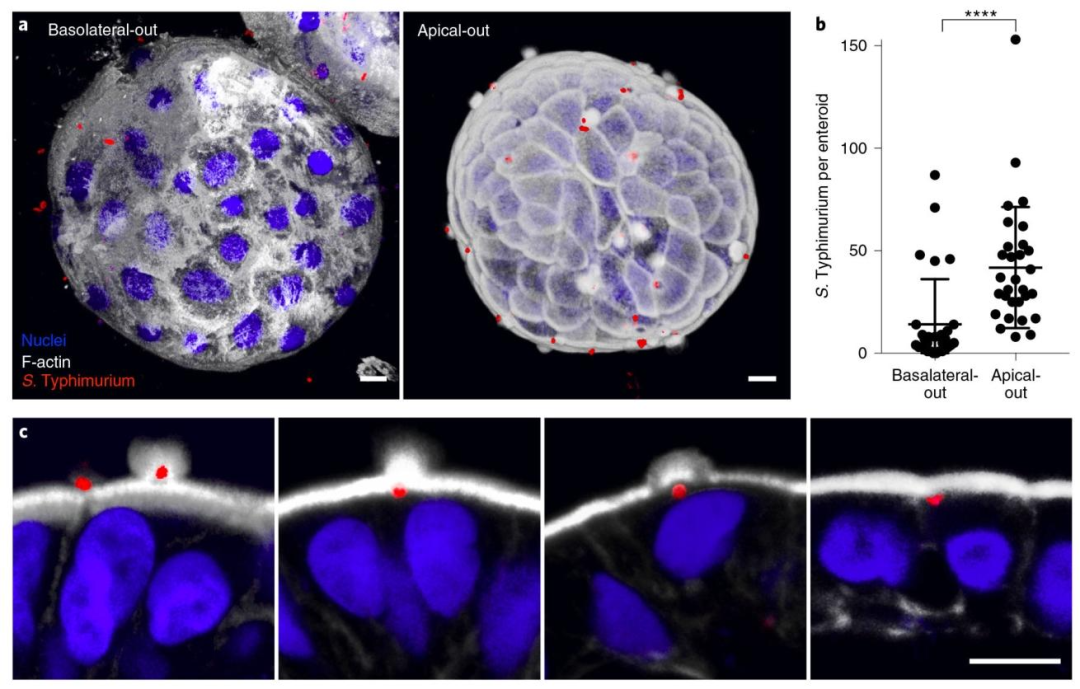
图 4:沙门氏菌感染顶 - 外肠类器官的实验验证
该图探究病原体感染的极性偏好性,(A)感染成像显示顶 - 内类器官中沙门氏菌(mCherry 标记,红)感染量少,顶 - 外类器官中细菌大量附着并入侵,F-actin(白)围绕细菌形成 “ruffles”(细胞骨架重排);(B)定量分析显示 1 小时感染后,顶 - 外类器官内细菌数(均值 120 个 / 类器官)显著高于顶 - 内类器官(均值 30 个 / 类器官,****P<0.0001,数据来自 3 次实验共 60 个类器官);(C)入侵细节显示沙门氏菌在顶膜表面聚集并诱导局部 F-actin 富集,证明沙门氏菌优先通过顶膜入侵胃肠道上皮,顶 - 外类器官是研究此类感染的理想模型。
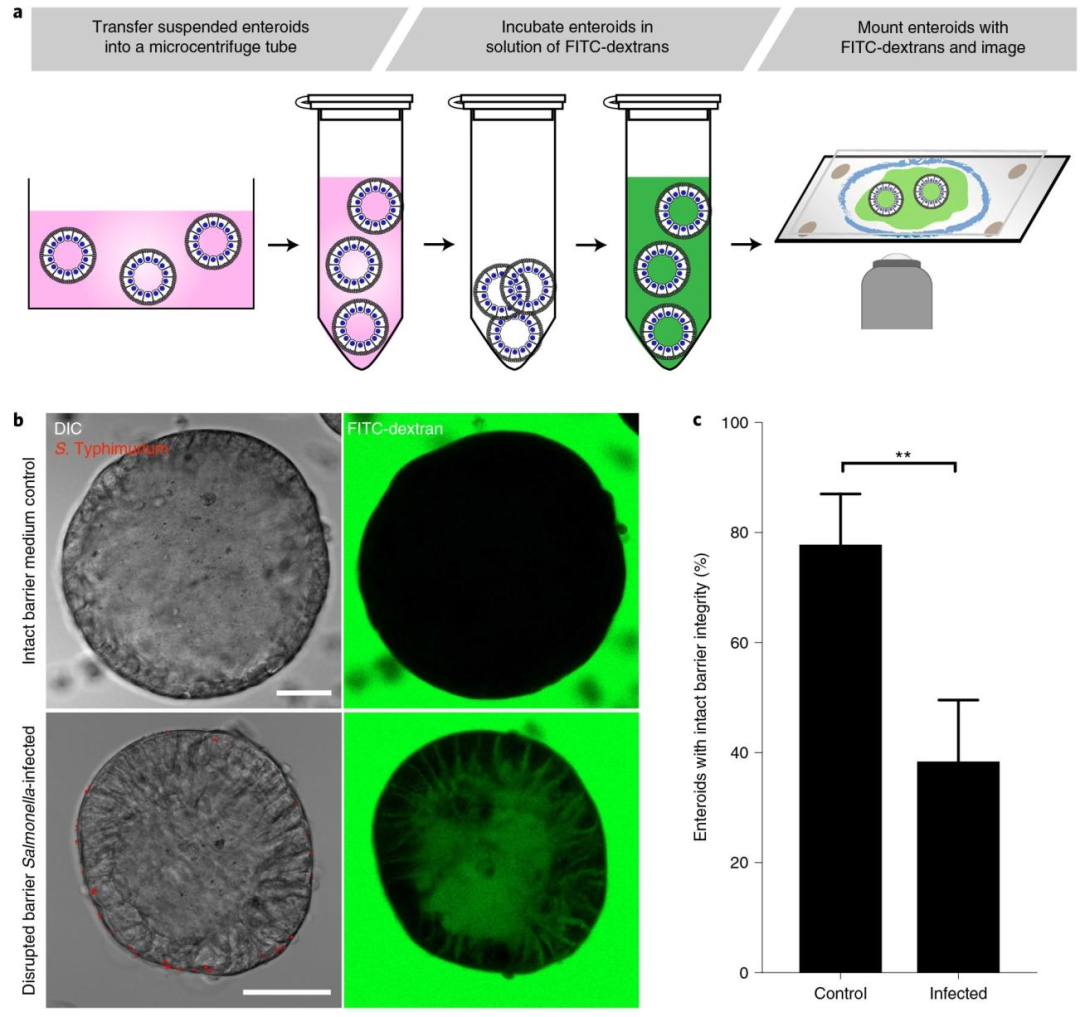
图 5:FITC - 葡聚糖扩散实验检测上皮屏障功能
该图建立顶 - 外类器官的屏障功能评估方法,(A)实验流程显示悬浮顶 - 外类器官与 4kDa FITC - 葡聚糖共孵育后立即活细胞成像,完整屏障排除葡聚糖,破损屏障允许葡聚糖渗入;(B)成像结果显示未感染(对照)顶 - 外类器官无荧光渗入,沙门氏菌感染 1 小时后荧光大量渗入;(C)定量分析显示对照类器官屏障完整率 82%±5%,感染组降至 28%±4%(P<0.01,数据来自 5 次实验共 369 个类器官),证明顶 - 外类器官可定量评估上皮屏障功能及病原体感染对屏障的破坏作用。
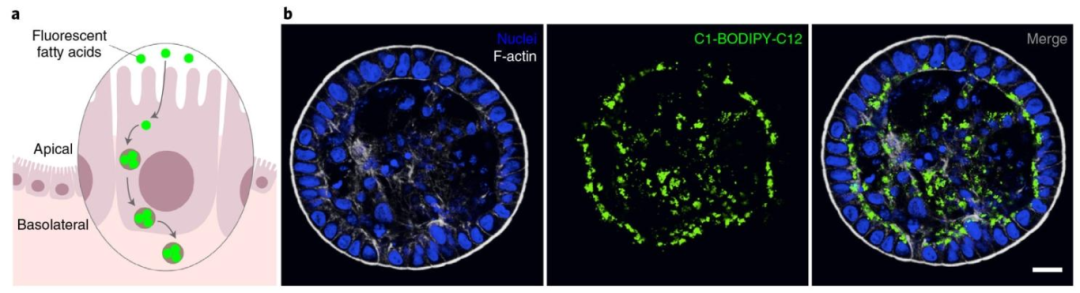
图 6:荧光脂肪酸吸收实验检测营养吸收功能
该图验证顶 - 外类器官的生理吸收能力,(A)实验示意图显示顶 - 外类器官的顶膜从培养基摄取荧光脂肪酸(C1-BODIPY-C12),吸收后形成细胞内脂滴并定位于基底侧;(B)成像结果显示 30 分钟孵育后,脂肪酸荧光(绿)以点状脂滴形式分布于细胞内且集中在基底侧(远离 F-actin 标记的顶膜,白),DAPI(蓝)标记细胞核,证明顶 - 外类器官保留正常的极性营养吸收功能,符合体内脂肪酸吸收后的转运规律。
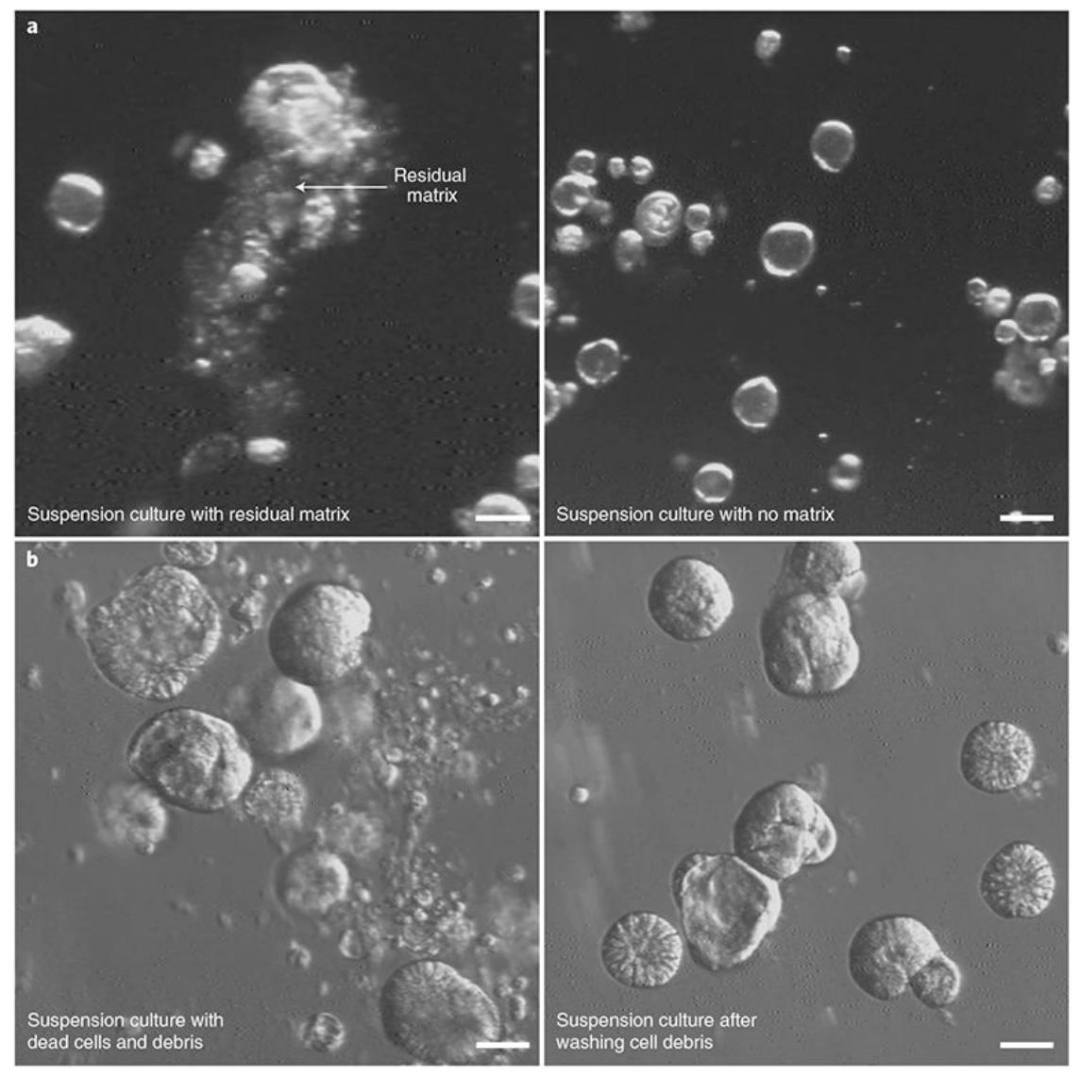
图 7:极性反转实验的常见问题与解决方案
该图针对实验操作关键问题提供优化策略,(A)残留基质问题显示左侧因 EDTA 处理不充分,残留 BME 导致类器官聚集、无法反转,右侧完全去除 BME 后类器官分散且成功反转;(B)细胞碎片问题显示左侧悬浮培养 1 天后类器官周围有大量死细胞与碎片,右侧通过离心洗涤去除碎片后类器官形态清晰,证明充分解离 BME 和增加洗涤步骤可解决聚集与碎片问题,提升实验重复性。
本研究针对胃肠道类器官顶膜难访问的核心问题,开发了高效可重复的极性反转方案:通过 EDTA 解离 BME + 悬浮培养,24-48 小时实现顶 - 外极性转化,保留上皮屏障完整性、多谱系分化及生理功能。通过 7 个核心 Figure 系统验证了方法的可行性(图 1-2)、调控规律(图 3)、应用价值(图 4-6)及操作优化(图 7),为感染性疾病、上皮功能研究提供关键工具。局限性在于顶 - 外类器官增殖较慢(不适用于长期传代),需使用未高度分化的类器官;未来可拓展至肺、肝等上皮组织,推动更多疾病模型的建立。
常见细胞污染类型如何辨别及预防解决方法:细胞培养中常见的生物污染类型有7种,分别是细菌污染,支原体污染,原虫污染,黑胶虫污染,真菌污染,病毒污染以及非细胞污染,真菌污染来源,一般是来自实验服,并且具有气候性,多雨······
细胞聚团的原因分析及如何避免:培养物中细胞可能聚集的一些原因包括:1.过度消化、2.环境压力、3.组织分解、4.过度生长、5.污染等;如何避免聚团细胞的生成;首先确认当前细胞生长密度及状态,80%左右的生长密度即可进行······
细胞有空泡原因分析及解决方法:出现细胞空泡情况有1.细胞老化2.培养液错误配制;3.细胞消化时操作不当;4.污染等等,如细胞老化,解决方法,原代细胞使用较低代次进行实验,传代细胞避免传代次数过高···
细胞半换液和全换液操作步骤:第一种方式:细胞全换液;如果是贴壁细胞,可以用全量换液法,直接吸去全部旧培养基,补充足量新鲜完全培养基;第二种方式:细胞半换液;"细胞半换液"又称"细胞半量换液",即弃掉一半旧的培养基,再······
细胞生长缓慢的可能原因有哪些:细胞培养外部因素包括细胞培养基的配方和质量问题,培养条件不理想,污染问题,细胞自身因素包含细胞的健康状态,细胞密度过高或过低,细胞老化现象,细胞特性,当细胞生长出现缓慢的问题时,我······
常用胰腺癌细胞株动物模型及胰腺癌细胞株有哪些:胰腺癌研究中常用的动物模型主要包括化学物质诱导胰腺癌动物模型,基因工程胰腺癌小鼠模型和胰腺癌移植模型,常用的胰腺细胞株MIA-PACA-2人胰腺癌细胞,Capan-2人胰腺癌细······
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
产品规格:1*10^6
¥3000
上一篇:人多能干细胞衍生视网膜类器官的高效可重复分化方法及视网膜命运特化机制解析
下一篇:基于诱导性 ETV2-iPSC 的肾脏类器官血管化构建及其在足细胞成熟与肾素细胞生成中的应用
厦门爱恪信生物科技有限公司
手机:15859239971
邮箱:2205839769@qq.com
地址:厦门翔安火炬高新区翔星路96号建业楼D座602

微信公众号
ATCC细胞培养

技术支持
15859239971
Copyright©厦门爱恪信 闽ICP备19027235号-7
公安备案: XML地图
XML地图




 客服QQ
客服QQ